

肥胖是重要公共卫生问题,影响着全球近20亿人口[1]。预计2030年中国将有70%以上的成年人和30%的儿童存在超重或肥胖问题,因超重或肥胖所引发的医疗费用高达4 180亿元,占全国医疗总费用的21.5%[2]。超重和肥胖是诱发高血压、Ⅱ型糖尿病(diabetes mellitus type 2, T2DM)、非酒精性脂肪肝(nonalcoholic fatty liver disease, NAFLD)、心血管疾病(cardiovascular disease, CVD)以及特定癌症(如乳腺癌、结肠直肠癌、肝癌、胰腺癌)等多种代谢疾病的重要危险因素[3-7]。中国成人的肥胖率在过去三十年间显著上升,从1982年的不到1%增长到2018年的51.2%,且这种趋势将持续增长[8]。中国儿童和青少年的肥胖率从2002年的6.6%急剧增长至2020年的20%,儿童及青少年肥胖不仅会影响大脑发育和心理健康,且该影响可能会一直持续到成年期,进而引发多种生理、心理和社会问题[9]。
肥胖或超重是由于能量摄入过多、消耗较少、脂肪代谢异常、体内脂肪异常堆积而引起的[8]。体育活动能够通过增加身体代谢率,促进脂肪氧化,减少体内脂肪堆积,维持肌肉质量和减少食欲来控制能量摄入,有效改善肥胖诱发的多器官代谢功能紊乱,继而提高人体免疫力,减轻氧化应激及慢性炎症,降低T2DM、NAFLD、CVD等多种代谢疾病的发病风险[10]。运动促进脂肪酸代谢的信号传导机制可能是通过脂肪酸诱导的氧化型辅酶Ⅰ(nicotinamide adenine dinucleotide,NAD+)刺激的沉寂信息调节因子(sirtuin 1,SIRT1)激活和/或脂肪酸介导的AMP活化蛋白激酶(AMP-activated protein kinase, AMPK)激活,直接或间接靶向过氧化物酶体增殖物激活受体(peroxisome proliferators-activated receptors,PPARs)的表达,促进脂代谢相关基因的表达,从而加速糖和脂肪酸氧化速率、线粒体生物合成及Ca2+重新摄取进入肌浆网[11]。然而,日常食物和饮料中糖类(如果糖、葡萄糖和麦芽糖浆等)的摄入对机体各器官功能会产生负面效应[12-14]。据报道,美国人均糖的摄取量相比于1970年增加了2倍以上,而在中国,每年人均糖的摄取量也在逐步增加[15]。青少年是含糖饮料和高糖精加工零食的主要摄入群体,这些含糖食品的摄入是导致肥胖的重要因素。“管住嘴,迈开腿”是预防和治疗肥胖和超重,降低相关代谢性疾病发病率,提升人民生活质量的主要策略。
高糖饮食与肥胖和多种慢性代谢性疾病的发生密切相关。研究表明,长期大量摄入含糖饮料和高糖食品与体重增加、腹部脂肪积累以及慢性疾病风险加剧之间显著相关[16]。高糖饮食不仅与肥胖、血脂异常和CVD发病风险增加正相关[17],还通过影响血管内皮功能、肠道屏障等途径诱发及恶化动脉粥样硬化[18]。大量摄入含有果糖的添加糖,如蔗糖和高果糖玉米糖浆,与糖尿病的发展密切相关[19]。
为降低糖摄入量,当前常用的饮食策略是在食品和饮料中使用热量较低或没有热量的甜味剂替代蔗糖、果糖和果葡萄糖浆等具有较高热量的添加糖类。近几年,含有甜味剂的“0”蔗糖食物和饮料广受运动人群和超重、肥胖及有减肥需求人群的关注,销售量逐年激增[20-21]。据报道,在减肥维持成功者中,低热量甜味剂的消耗量远高于一般成年人群,在成功保持体重减轻大于一年的个体中,约有66%的人报告每周饮用含甜味剂的饮料,包括无糖软饮料、含甜味剂的咖啡/茶/果汁、运动饮料等[22-23]。
值得注意的是,尽管甜味剂不具有或具有较低的热量,但其是否能够有效预防体重增加并阻断肥胖发生目前尚具有争议性[24-26]。甜味剂可以被人体吸收并影响机体重要代谢器官如肝脏、脂肪组织,或作用于肠道微生物并对菌群代谢产生显著影响[27-29]。鉴于运动与甜味剂摄入在肥胖发生和发展中的关键作用,本文通过检索中国知网、PubMed、Web of Science等数据库,详细梳理运动通过多器官、多靶点影响肥胖及相关代谢疾病的关键作用机制,总结不同类别甜味剂对肥胖及相关代谢疾病关键蛋白和基因靶点的影响,揭示甜味剂摄入对运动改善肥胖及其相关代谢疾病健康作用的潜在协同或拮抗作用,为肥胖等慢性疾病的运动预防和康复提供理论依据和实践指导。
1 甜味剂摄入对肥胖及其相关代谢疾病发生和发展的影响
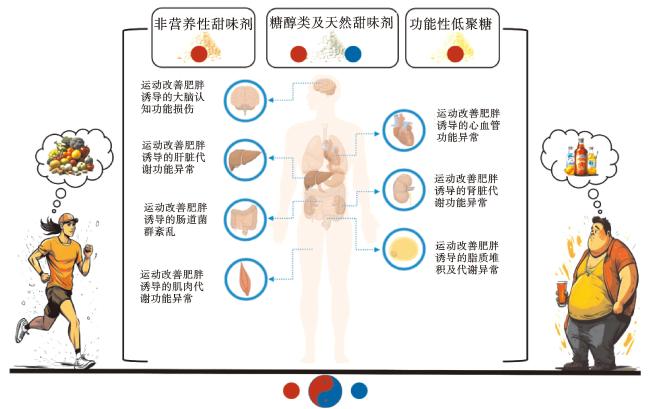
含有甜味剂的“0”糖食物和饮料受到广大减肥、健身以及糖尿病人群的推崇,被认为是糖的安全替代品。目前,主要用于食品加工和临床研究的甜味剂主要有3类:非营养性甜味剂,又称人造甜味剂,主要包括阿斯巴甜、糖精、纽甜、甜蜜素和三氯蔗糖,这些甜味剂几乎不含能量且甜度很高;糖醇类及天然非营养性甜味剂,包括赤藓糖醇、阿洛酮糖、甜叶菊糖苷和罗汉果提取物等;功能性低聚糖,主要包括植物来源的功能性低聚糖(如大豆低聚糖)、微生物发酵产生的低聚糖(如低聚异麦芽糖)以及人工合成的低聚糖(如低聚木糖)。这些功能性低聚糖都具有良好的生物活性,例如抗氧化、抗菌抑菌、降血脂、降血糖、增殖肠道益生菌等。
关于甜味剂是否有利于减少肥胖、改善肥胖相关代谢疾病的研究结果存在不一致性。一项针对超重成年人(n=303)的干预性研究发现,与每天饮用710 mL饮料的人相比,摄入同样多的甜味剂饮料(每237 mL热量小于5 kcal)有助于减肥和维持体重[30]。然而,Higgins等[31]基于154名健康受试者开展的随机临床试验表明,每天饮用1.25~1.75 L含有糖精、阿斯巴甜的饮料与肥胖发生风险增加显著正相关。通过对10万名50~79岁绝经女性开展长达21年的随访研究发现,与每月饮用3份(每份355 mL)及以下的含糖饮料相比,每天饮用1份或更多含糖饮料(糖精、三氯蔗糖等)的绝经女性患肝癌风险增加85%,死于慢性肝病的风险增加68%[13]。
使用甜味剂代替食品中的添加糖可降低热量摄入,但尚无充分证据表明糖尿病患者摄入甜味剂,相较于糖、安慰剂或其他营养性低热量替代品,在改善血红蛋白A1c(hemoglobin A1c, HbA1c)水平、控制肥胖以及减少不良事件方面存在显著优势或危害[32]。人工甜味剂(如三氯蔗糖、阿斯巴甜、糖精等)会削弱机体肠道对葡萄糖的吸收能力,降低胰岛素和肠促胰岛素的分泌,改变肠道微生物群结构及功能,恶化糖尿病患者的血糖控制[33]。糖精也能够抑制对糖的味觉反应,增强胰岛素分泌[34]。相反,甜菊糖苷和甜菊醇可以调控β细胞,刺激胰岛素分泌,改善机体糖脂代谢和肠道微生态,调节核因子κB(nuclear factor kappa-B, NF-κB)等炎症和糖脂代谢关键基因的表达,对抑制T2DM有一定积极作用[35]。
一项基于NutriNet-Santé队列(n=103 388)的前瞻性研究表明,人工甜味剂摄入(特别是阿斯巴甜、安赛蜜和三氯蔗糖)与CVD风险之间存在直接联系[36]。基于Discovery队列(n=1 157)、US validation队列(n=2 149)及European validation队列(n=833)开展的研究发现,赤藓糖醇的循环水平与重大不良心血管事件,包括复发心绞痛、急性心肌梗死、严重心律失常、心力衰竭、冠心病死亡、心肌梗死或中风风险密切相关,并会引发血小板聚集和血栓形成[14]。Pacheco等[37]对105 148名参与者调查发现,非营养性甜味剂与心肌梗死、急性冠状动脉综合征等CVD高风险相关,高摄入量甜味剂组(平均摄入176.7 mL/d)首次发生心血管事件的风险更高。相反的是,甜菊糖苷可能会通过降血压、降血糖、抗氧化、抗炎等途径起到保护心血管的作用[38]。一项随机双盲对照试验中,每天3次服用含有500 mg甜菊苷粉的胶囊或安慰剂,持续2年后,甜菊苷组人群的平均收缩压和舒张压显著下降[39]。
综上所述,甜味剂对糖尿病及其相关代谢异常的影响是多方面的,涉及能量代谢、胰岛素敏感性、炎症反应等多个生理环节。未来研究需要进一步探索甜味剂摄入的剂量、类型及其与生活方式(包括饮食模式、运动等)的相互作用,以更好地理解甜味剂对健康的影响,指导公众做出更加健康的选择。
2 甜味剂对运动改善肥胖诱发的多器官代谢异常的协同/拮抗作用
运动通过多途径、多层次作用改善肥胖引起的多器官代谢异常,包括胰岛素信号传导的优化、糖脂代谢调控因子的调节、脂肪组织表型的改变、炎症与氧化应激的调节、瘦素信号的改善以及趋化素调控等(图1)。甜味剂已被广泛应用于食品加工和临床研究中,且甜味剂摄入与机体能量代谢、脂肪代谢等关系复杂,提示甜味剂摄入或可对运动改善肥胖及其相关代谢疾病健康作用存在潜在协同或拮抗作用,然而现阶段尚无对二者潜在交互作用的关系探究,亟待开展人群或动物实验,深入研究二者是否存在共同分子机制。鉴于运动与甜味剂摄入对肥胖发生和发展关键靶点的调节作用,本文梳理二者对不同靶点的组织特异性调节作用(图2),对食-体联合防治肥胖等慢性疾病研究有重要指导意义。
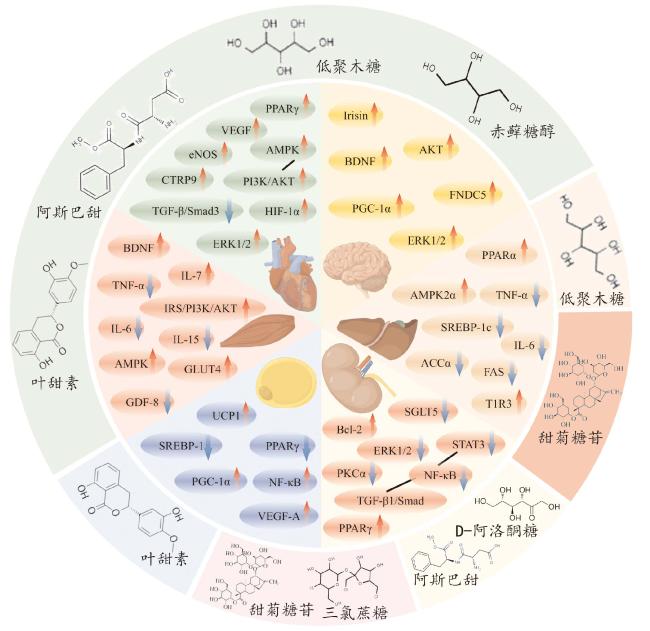
图1 不同甜味剂对运动组织特异性靶点改善作用的协同与拮抗作用注:图中外圈不同颜色代表作用于不同运动组织的共同甜味剂;内圈扇形代表各组织中甜味剂或运动共同作用的靶点。箭头表示甜味剂或运动对组织特异性靶点的影响,“↑”表示激活或增加,“↓”表示抑制或减少,蓝色箭头代表协同作用,红色箭头代表拮抗作用。网络版为彩图。 Fig.1 Synergistic and antagonistic effects of different sweeteners on the improvement of exercise-induced tissue-specific targets |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
2.1 甜味剂对运动改善脂肪功能靶点的影响
肥胖导致脂肪细胞激素(如瘦素、脂联素、抵抗素)分泌失调、脂肪细胞沉积和脂代谢异常,降低脂肪细胞对胰岛素刺激的敏感性,影响脂肪分解及转运,造成脂肪异常积累[40]。运动可通过激活线粒体解偶联蛋白1(uncoupling protein 1, UCP1)、血管内皮生长因子A(vascular endothelial growth factor-A, VEGF-A)、过氧化物酶体增殖物激活受体-γ共激活因子-1α(peroxisome proliferator-activated receptor γ coactivator-1α, PGC-1α)促进棕色脂肪组织产热和白色脂肪组织米色化[41-44],改善脂肪代谢。
叶甜素是一种在绣球花变种中发现的天然甜味剂。它可以通过上调UCP1和PGC-1α,同时下调过氧化物酶体增殖物激活受体γ(peroxisome proliferator-activated receptor-gamma, PPARγ)和固醇调节元件结合蛋白-1c(sterol regulatory element binding protein-1c, SREBP-1c),改善高脂饮食诱导的肥胖小鼠相关基因表达,以减少皮下脂肪、瘦素水平和代谢异常,包括与血糖和脂质相关的异常[48]。饲喂甜菊糖苷的肥胖大鼠脂肪细胞PPARγ表达降低,UCP1表达显著增加[49]。相反,人工甜味剂三氯蔗糖摄入可显著升高大鼠炎症因子NF-κB的表达,抑制PPARγ的表达,促进脂肪细胞肥大,进而加剧脂质积累,这或与运动改善肥胖诱发的脂肪功能发生拮抗作用[50]。
2.2 甜味剂对运动改善骨骼肌功能靶点的影响
肥胖会减少骨骼肌质量,改变骨骼肌纤维类型,降低肌肉线粒体数量和活性,引发运动力量和耐力下降,诱发肌肉萎缩、骨质疏松症以及机体糖脂代谢失衡等疾病[51-52]。运动改善机体肥胖得益于骨骼肌分泌的运动因子。多项证据表明,肥胖与生长分化因子8(growth differentiation factor-8, GDF-8)表达增加有关,有氧运动和力量运动会减弱GDF-8的表达,增强耐力运动对代谢的有益作用[53-54]。运动通过肌动蛋白的自分泌/旁分泌机制调节葡萄糖摄取,刺激脑源性神经营养因子(brain-derived neurotrophic factor, BDNF)和白细胞介素-6(interleukin-6, IL-6)参与AMPK介导的脂肪酸氧化,调控GDF-8、白血病抑制因子(leukemia inhibitory factor, LIF)、IL-6、白细胞介素-7(interleukin-7, IL-7)和白细胞介素-15(interleukin-15, IL-15)参与肌肉肥大和肌生成,改善高脂饮食诱发的骨骼肌功能障碍[55-56]。此外,有氧运动通过上调IRS-1/PI3K/AKT信号通路,抑制NF-κB的激活,减轻高脂饮食肥胖小鼠骨骼肌炎症,进而改善胰岛素抵抗和葡萄糖代谢[57]。
2.3 甜味剂对运动改善肝脏功能靶点的影响
肥胖导致肝脏胰岛素抵抗、瘦素抵抗和脂质代谢紊乱,脂肪堆积在肝脏中,引发NAFLD等相关疾病,甚至可能发展为肝炎、肝硬化、肝癌等[61-62]。运动可有效降低肥胖大鼠体重,其通过上调成纤维细胞生长因子21(fibroblast growth factor 21, FGF-21)和PGC-1α,促进脂肪酸氧化,改善脂代谢[63];也可激活肝脏磷脂酰肌醇3激酶(phosphatidylinositol 3 kinase, PI3K)磷酸化,减少肝内瘦素分泌,提高瘦素受体表达,减少肥胖引起的肝脏脂肪积累[64-65]。有氧运动还通过上调AMPK和过氧化物酶体增殖物激活受体α(peroxisome proliferator-activated receptor-alpha, PPARα)蛋白表达,降低大鼠IL-6、TNF-α、白细胞介素10(interleukin-10, IL-10)表达,减缓肥胖大鼠的肝脏脂肪变性、炎症和胰岛素抵抗[66];或可通过抑制肝脏α-1,3-甘露糖糖蛋白2-β-N-乙酰氨基葡萄糖转移酶(alpha-1,3-mannosyl-glycoprotein 2-beta-N-acetylglucosaminyltransferase, MGAT1)改善脂代谢[67]。
研究表明,高果糖摄入会刺激肝脏脂质新生并增加脂质分泌和胆固醇水平[68]。相反,甜菊糖苷摄入可降低肥胖大鼠总胆固醇(total cholesterol, TC)、甘油三酯(triglyceride, TG)和低密度脂蛋白(low-density lipoprotein, LDL)水平,升高高密度脂蛋白(high-density lipoprotein, HDL)水平,降低大鼠肝脏促炎因子,如白细胞介素1β(interleukin-1β, IL-1β)、IL-6和TNF-α,改善肝脏氧化应激[69]。大鼠腹腔注射甜菊糖苷(200 mg/kg)还可以通过调节核红细胞因子2(nuclear factor-erythroid 2-related factor 2, Nrf2)的水平来预防肝损伤[70]。D-阿洛酮糖[71]、低聚果糖和低聚半乳糖[72]可以加速脂质消耗,抑制脂肪生成相关因子乙酰辅酶A羧化酶α(acetyl-CoA carboxylase, ACCα)、肝脏脂肪酸合成酶(fatty acid synthase, FAS)、SREBP-1c的表达,同时刺激脂肪酸氧化相关因子AMPK、激素敏感性脂肪酶 (hormone-sensitive triglyceride lipase,HSL)和PPARα表达,减少炎症相关标志物,改善肝脏脂质代谢。然而,最新研究发现,高剂量D-阿洛酮糖(0.75 mg/mL)会降低肠道菌群的α-多样性和短链脂肪酸含量,升高厚壁菌门/拟杆菌门的比例,损伤结肠黏膜屏障,进而恶化葡聚糖硫酸钠(dextran sulfate sodium, DSS)诱导的小鼠结肠炎[73]。三氯蔗糖可能激活同源甜味受体(taste receptor type1 member 3, T1R3),产生活性氧(reactive oxygen species, ROS),促进内质网应激和脂肪生成,加剧高脂饮食诱导的肝脂肪变性[50]。摄入阿斯巴甜可能通过“肝-肠轴”增加肠道内梭状芽孢杆菌的丰度,诱导胰岛素抵抗,对肝脏产生损伤[74]。
2.4 甜味剂对运动改善肾脏功能靶点的影响
肥胖会导致肾功能损伤,引发肾小球病变、糖尿病肾病、高血压肾病等肾脏疾病,促进慢性肾病(chronic kidney disease, CKD)的发生[75]。运动能够提升CKD患者的最大心率并降低静息心率,增加患者透析前的肌肉力量[76]。定期运动可以降低血浆IL-10水平,下调T淋巴细胞和单核细胞,通过促进蛋白质合成、增加肌肉线粒体含量和减少肌肉分解代谢,对抗CKD诱发的肌肉萎缩[77]。运动还可以通过调节参与线粒体凋亡途径的转化生长因子-β1(transforming growth factor-β1, TGF-β1)和B淋巴细胞瘤-2(B-cell lymphoma-2, Bcl-2)蛋白表达,减轻肾间质纤维化,改善高血压引发的肾功能损伤[78]。
相反,Shen等[82]研究发现甜菊糖苷对小鼠肾纤维化具有抑制作用,其主要机制可能与PPARγ的激活有关,并通过下调NF-κB介导的TGF-β1和信号传导及转录激活蛋白3(signal transducer and activator of transcription 3, STAT3)表达,阻断Smad介导的纤维化信号通路。甜菊糖苷还通过调节Bcl-2、细胞外调节蛋白激酶1/2(extracellular regulated kinase 1/2,Erk1/2)蛋白表达,减轻肾脏细胞凋亡和氧化应激[83]。肥胖和糖尿病患者肾脏有机阴离子转运体3多肽B(organic anion transporter 3 polypeptide B, Oat3)功能受损,而低聚木糖干预可以改善肠道菌群失调,增加肠道微生物群中有益菌(双歧杆菌属、嗜粘液阿克曼氏菌和普氏粪杆菌)的相对丰度,减少高脂血症和低度全身炎症,改善胰岛素敏感性,从而恢复肾脏Oat3的功能[84]。
2.5 甜味剂对运动改善脑功能靶点的影响
超重或肥胖会严重影响大脑活动,增加罹患阿尔茨海默病以及许多其他精神和认知疾病的风险[85]。运动可以激活大脑新陈代谢,增加脑血流量,刺激神经递质分泌,进而对大脑功能产生积极影响。研究表明,机械振动可以激活老年痴呆患者大脑感觉(运动)皮层,诱导小脑、丘脑等区域的血流增加和突触增强,刺激肌梭和感觉器官,增加脑电活跃度,改善老年痴呆患者认知功能和身体平衡机能[86]。BDNF对促进中枢神经系统的生长和健康至关重要,也是胃肠道紧密连接蛋白表达和调节的重要因子。运动通过PGC-1α/纤连蛋白Ⅲ型结构域蛋白5(fibronectin type Ⅲ domain-containing 5, FNDC5)信号通路诱导海马BDNF和其他神经保护因子的表达,激活鸢尾素(irisin)/BDNF通路,改善突触可塑性和认知功能[87-89]。Li等[90]研究发现,鸢尾素通过激活蛋白激酶B(protein kinase B, Akt)和Erk1/2信号通路减少缺血诱导的神经元损伤,有助于运动对脑缺血的神经保护作用。
高果糖和高蔗糖饮食摄入会导致更强烈的饥饿感和对食物的渴望,促使三磷酸腺苷(adenosine triphosphate, ATP)快速消耗,降低下丘脑中的丙二酰辅酶A(malonyl-CoA)水平,从而增加食物摄入量[68]。研究发现,长期摄入赤藓糖醇增加肠道胆囊收缩素(cholecystokinin, CCK)和胰高血糖素样肽-1(glucagon-like peptide-1, GLP-1)释放,并加重小鼠脑缺血损伤[91-92]。雄性C57BL/6J小鼠长期摄入安赛蜜会导致神经元代谢改变和认知功能障碍,这可能与BDNF和Akt/Erk介导的细胞生长/存活途径失调有关[93]。在孕期摄入阿斯巴甜会增加食欲,导致肥胖饮食,对雌鼠及其子代体重管理、血糖控制和新陈代谢产生不利影响[12],即便每日摄入8%~15%的规定食用量,阿斯巴甜仍可影响γ-氨基丁酸分泌,增加焦虑情绪甚至子代抑郁的风险[94]。与之相反的是,功能性低聚糖干预可通过“肠-脑轴”改善自闭症患者的血清素水平和多巴胺代谢紊乱[95]以及阿尔兹海默症小鼠的认知功能障碍[96]。最近研究表明,叶甜素可以改善高脂饮食导致的BDNF减少,减轻 CD-1小鼠β-淀粉样蛋白(amyloid β-protein,Aβ)诱导的记忆障碍,并减少小鼠海马体的Aβ 聚集,改善突触可塑性和神经炎症。
综上,长期摄入人工甜味剂(阿斯巴甜)和糖醇类甜味剂(赤藓糖醇)可能通过BDNF/Akt/Erk信号通路与运动产生拮抗作用,加剧肥胖诱发的大脑认知功能障碍。功能性低聚糖和叶甜素可能通过“肠-脑轴”促进BDNF释放,与运动协同减少神经炎症并改善突触可塑性。
2.6 甜味剂对运动改善心血管功能靶点的影响
肥胖是冠心病、高血压等心血管疾病的重要诱因。运动可有效降低CVD风险,改善心功能。抗阻运动能够显著提高肥胖大鼠心肌去乙酰化酶3(sirtuin 3, SIRT3)表达,调控心肌线粒体长链脂酰辅酶A脱氢酶(long-chain acyl-Co A dehydrogenase, LCAD)和b-羟基酰基-CoA脱氢酶(b-hydroxyacyl-Co A dehydrogenase, b-HAD)的活性,改善高脂饮食诱导的肥胖心功能障碍[97]。运动还可以显著增加血清和心肌补体1q/肿瘤坏死因子相关蛋白9(C1q/tumor necrosis factor related proteins9, CTRP9)水平,激活AMPK、PI3K/Akt信号通路,促进蛋白质合成,减少心梗模型小鼠蛋白质降解和细胞凋亡,降低血压[78,98 -99]。研究揭示,运动通过增加小鼠心脏内皮型一氧化氮合酶(endothelial nitric oxide synthase, eNOS)、VEGF和缺氧诱导因子-1α(hypoxia-inducible factor-1α, HIF-1α)表达,促进糖尿病小鼠心肌血管生成,改善胰岛素抵抗和血管内皮功能障碍[100]。运动训练通过调节Erk1/2、TGF-β/Smad3等信号通路,缓解炎症与氧化应激水平,改善心脑血管结构与功能,发挥对心脑血管系统的保护效应[101]。
高糖饮食会影响血管内皮活性,增加肠道通透性,提高循环中细胞因子水平,促进动脉粥样硬化、高血压的发生发展。甜菊糖苷在临床研究中被描述为血管扩张剂,其通过抑制钙离子内流来降低血管阻力,起到保护心血管的作用[102]。Wang等[103]研究发现,小鼠连续摄入0.2 mg甜菊糖苷40 d后,其心肌组织中TGF-β1、NF-κB、Smad2/3表达显著降低, PPARγ、Smad7表达增加,表明甜菊糖苷可以对小鼠心肌纤维化发挥保护作用。经甜菊糖苷处理后,大鼠成纤维细胞中的谷胱甘肽(glutathione, GSH)水平以及超氧化物歧化酶(superoxide dismutase, SOD)的表达和活性显著增强,并通过激活 PI3K/Akt 通路导致大鼠成纤维细胞葡萄糖摄取增加,从而诱导GLUT4易位到质膜[104]。
3 结论与展望
运动是一种健康和低成本的减重方式,对肥胖相关代谢性疾病的预防与康复具有积极作用。运动通过减少脂肪尤其是内脏脂肪的堆积,提高能量消耗以及胰岛素敏感性,改善脂肪组织、心脑血管、肝脏、骨骼肌、肠道等多器官代谢功能。众所周知,良好的饮食习惯对健康有诸多益处,“管住嘴,迈开腿”不仅可以有效管理体重,也是提升人民生活质量和增长寿命的健康策略。“控糖、减糖”的减重食谱推动低热量甜味剂在食品加工行业的快速发展。然而,甜味剂与健康之间的关系较为复杂。甜味剂不具有或具有较低的热量,但是不同种类的甜味剂在体内的吸收代谢途径不同,个体差异性大,其是否能够有效预防体重增加并阻断肥胖发生目前仍然具有争议。此外,现有关于甜味剂的研究主要集中于单一甜味剂产生的效应,而大多数市面上销售的产品中添加了2种以上的复合甜味剂,甜味剂混合暴露对人体的健康效应尚不明确。
不同种类的甜味剂对运动改善肥胖诱发的多器官功能代谢异常具有不同的作用。人工甜味剂(例如阿斯巴甜、安赛蜜、三氯蔗糖等)和糖醇类甜味剂(如赤藓糖醇)可能通过调控PGC-1α/FNDC5、PI3K/Akt/Erk等信号通路,刺激BDNF、NF-κB、IL-6、irisin等因子释放,参与运动产生的拮抗作用,进而降低认知功能,加剧神经炎症,并损害肝脏、心血管、骨骼肌和脂肪等组织的功能。相反,天然甜味剂和功能性低聚糖(例如甜菊糖醇、低聚木糖、叶甜素等)则可能通过调控IRS/PI3K/Akt、AMPK/ACC、PPAR等通路介导的脂肪酸氧化,与运动协同下调NF-κB介导的IL-6、TNF-α等炎性因子表达,阻断Smad纤维化信号通路,参与维持血糖稳态平衡、脂肪酸氧化、线粒体功能,从而改善肥胖及其相关的代谢紊乱。尽管本文总结了不同类别甜味剂对运动改善肥胖作用关键靶点的影响,并揭示了甜味剂摄入对运动改善肥胖及其相关代谢疾病健康作用的潜在协同或拮抗作用,但目前仍缺乏实验数据的验证。因此,亟需开展动物及人群的随机对照实验,以明确甜味剂摄入对运动健康效应的具体影响,从而更好地指导肥胖防控和健康生活实践。
甜味剂消费可能对体重管理产生影响,但这种影响受到多种因素的制约,例如个体代谢率、饮食习惯以及生活方式等。在补充甜味剂时,应将其与均衡饮食和适度运动科学地结合起来,以便达到最佳减重效果,改善代谢健康。