

细胞壁是植物细胞的特征之一,主要由多糖、酚类化合物及多种蛋白质等生物聚合物构成,这些成分赋予了细胞壁机械强度和刚度。细胞壁的结构特性不仅决定了植物细胞的性质,还直接影响到植物器官的发育及对环境刺激的响应[1]。植物细胞壁分为初生细胞壁和次生细胞壁,两者在组成成分和细胞位置上有所不同。植物细胞都具有较薄且可伸展的初生细胞壁,其在细胞分裂过程中合成,对细胞伸长和膨胀起关键调节作用,进而影响细胞形状和大小[2-3]。次生细胞壁较厚且坚硬,形成于特定细胞(如木质部细胞)中,赋予细胞额外的机械刚度和疏水性[4]。纤维素是初生细胞壁和次生细胞壁的主要成分,但其在初生细胞壁中聚合度较低,微纤维较窄;在次生细胞壁中聚合度高,微纤维较宽。此外,初生细胞壁主要含木葡聚糖和果胶,而次生细胞壁富含半纤维素、木质素和酚类聚合物。这些成分差异使得初生细胞壁具有柔韧性,次生细胞壁则在机械和生物学上更稳健。
在植物生物学的深入探索中,次生细胞壁的生物合成机制一直是研究重点。自2005年以来,通过多项系统性研究,已经确定多个在次生细胞壁生物合成过程中发挥关键调控作用的转录因子。例如,Kubo等[5]利用体外细胞培养技术,深入剖析了植物NAC转录因子家族,包括NAM、ATAF1、ATAF2、CUC2,并明确VND1-7在调控木质部导管细胞分化过程中的核心角色。Mitsuda等[6]发现了NAC家族次生壁增厚促进因子NST1和NST2,它们在花药细胞中调节次生细胞壁的形成。随后研究表明,NST1和NST3(又称次生壁相关NAC结构域蛋白1/SND1)在拟南芥中为调控纤维细胞分化的主效调控因子[7-8]。这些发现揭示了分子水平上对次生细胞壁生物合成的调节,表明植物具有调节次生细胞壁生物合成的特异性转录开关,这些因子属于NAC家族,包括VND和NST转录因子[9-11]。SND1和NST1在调控束间纤维次生细胞壁合成中发挥作用[12]。VND和NST/SND在多种维管植物中均具有同源性,表明它们在维管植物中普遍起着调节次生细胞壁发育的作用。在这些关于NAC蛋白的报道之后,另有研究表明多种MYB型转录因子是次生细胞壁形成的二级主调节因子[13-16]。MYB转录因子广泛存在于真核生物中,其N端具有高度特异性和保守性的DNA结合结构域,称为R1、R2、R3结构域。在植物中,大多数MYB蛋白只有2个结构域,因此被称为R2R3-MYB蛋白。有研究指出,MYB转录因子MYB46和MYB83在调控植物次生细胞壁合成的复杂网络中占据着至关重要的地位,它们充当这一调控过程中的二级关键控制点。在拟南芥中通过启动子活性分析和突变体表型观察,发现AtMYB46和AtMYB83在维管发育中发挥关键作用,特别是在束间纤维细胞分化调控中表现出显著活性。研究还发现,当MYB46和MYB83的表达水平降低时,植物次生细胞壁的增厚特征会丧失。进一步的实验结果显示,MYB46或MYB83的过表达能显著增强与次生细胞壁相关的下游基因表达,并促进纤维素、半纤维素、木质素积累。这些发现揭示了拟南芥次生细胞壁形成受NAC家族转录因子主导的多层级调控网络驱动,其中NAC主效调控因子通过激活次级MYB类调控因子,协同调控纤维素、半纤维素、木质素生物合成相关靶基因的时序性表达。
苦荞(Fagopyrum tataricum(L.)Gaertn.)通常称作鞑靼荞麦或三角麦,也被称为莜麦、乌麦、花荞等,是蓼科下的双子叶植物[17]。它主要种植于中国西南地区,以其独特的营养成分而闻名,包括黄酮类、酚酸类、三萜类、苯丙苷类、生物活性多糖类、膳食纤维、生物活性蛋白肽类、D-手性肌醇及其衍生物等多种具有功能的活性物质[18-19]。苦荞作为我国传统小杂粮作物和药食同源植物[20],被誉为“五谷之王”及“三降食品”[21-22]。它不仅具有养身、保健和食疗的三重功效,而且有助于强化肠胃功能,促进代谢废物排出。在现代食品加工领域,苦荞也被视为功能性谷物[23]。目前,在农业生产领域广泛种植的苦荞主要是果壳厚重且坚硬的品种。苦荞种子被高度木质化的果壳包裹,其致密的次生细胞壁结构导致脱壳效率显著降低且籽粒破损率增加。对苦荞果壳的细胞构造、生物学功能及其分子合成途径进行系统性探究,有利于培育易于脱壳的薄壳苦荞新品种,推动苦荞加工业高层次发展。陈庆富等[24]经过对比研究,发现薄壳苦荞与厚壳苦荞在果壳形态上存在显著差异,薄壳苦荞果壳腹面棱间区厚度明显低于厚壳苦荞相应部位的厚度。此外,薄壳苦荞在该区域缺乏沟槽结构,可能是导致其果壳易于脱落的形态学原因。Li等[25]对比分析了厚壳与薄壳苦荞在不同生长阶段的籽粒果壳细胞,生理、转录组和基因共表达研究发现,在薄壳苦荞的早期发育阶段,与次生壁合成相关的众多调控基因,如NAC转录因子和结构基因,表达水平较低,导致薄壳苦荞果壳中的纤维素和半纤维素含量明显低于厚壳苦荞,其果壳细胞次生壁的厚度显著减小。可见,薄壳苦荞的果壳更为薄弱,易于脱壳,而苦荞果壳细胞的次生壁合成在决定脱壳难易度上扮演着关键角色。
本课题组前期通过转录组分析,鉴定到1个苦荞果壳细胞次生壁生物合成候选NAC转录因子FtNAC16,其主要在苦荞纤维化程度较高的组织,如根、茎、花、发育的种壳中高表达,暗示其可能调控相关组织部位细胞次生壁的合成[26]。本研究在此基础上,利用分子生物学手段获得FtNAC16的过表达转基因毛状根并进行表型及表达分析,验证其生物学功能,为进一步剖析FtNAC16在苦荞果壳细胞次生壁生物合成中的功能及其分子调控机制提供参考。
1 材料与方法
1.1 实验材料
植物材料:厚壳苦荞‘品苦荞1号’来自贵州师范大学荞麦产业技术研究中心试验基地。
实验菌株:大肠杆菌(Escherichia coli)DH5α、Trans1-T1、发根农杆菌(Agrobacterium rhizogenes)Ar A4均来自本实验室。
载体:入门载体pDONR221(卡那霉素抗性)和过表达载体pK7WG2D(细菌中为壮观霉素抗性、植物中为卡那霉素抗性)均保存于本实验室。
实验所用引物如表1所示。
表1 本实验所用引物信息Tab.1 Primer information used in this study |
| 引物名称 | 引物序列(5'-3') | 用途 | ||
|---|---|---|---|---|
| FtNAC16-f1 | GCAGCCATGAATCTCTCAGTCAA | 基因克隆 | ||
| FtNAC16-r1 | CTCCAATCATACCGAGAAATGAGAG | |||
| FtNAC16-f过 | GGGGACAAGTTTGTACAAAAAAGCAGGCTCCGCAGCCAT GAATCTCTCAGTCA | 构建过表达载体 | ||
| FtNAC16-r过 | GGGGACCACTTTGTACAAGAAAGCTGGGTGCTCCAATCA TACCGAGAAATGAGAG | |||
| FtNAC16-qf | CTACTACTGCTACTGCTACG | 实时荧光定量PCR | ||
| FtNAC16-qr | GGAAGAGGGAGAAGAAGACT | |||
| FtMYB83-qf | ACAAGACATTACGGCGAGCT | |||
| FtMYB83-qr | GCACGAGGATAACGATGATG | |||
| FtMYB103-qf | GAAAATCGTCGTCATCCTCC | |||
| FtMYB103-qr | CGAACATGAAGAATGGAGCC | |||
| FtMYB54-qf | CCATTACTACGACTTCCTCC | |||
| FtMYB54-qr | ACCTTGTCTCGTTCTCGTTC | |||
| FtCESA4-qf | AGGTGGAGCAATGTAGGCAT | |||
| FtCESA4-qr | TCTGCTGCCTTTGATGTCAC | |||
| FtCESA7-qf | GTTGCTGGGATTTCAGATGC | |||
| FtCESA7-qr | GAGAAGATGGAAGCCAAGAG | |||
| FtCESA8-qf | TCTCGGCTCATCTCTTTGCA | |||
| FtCESA8-qr | GTCGGAGGAATAAGCAAGGT | |||
| FtIRX9-qf | GGCTGGCATTTGAAGAAGAC | |||
| FtIRX9-qr | TCCTGCTGTTTTGTGCCTTG | |||
| FtTBL3-qf | AGCATTGGGGTGATGTTGAC | |||
| FtTBL3-qr | AGTTTGCCCATGTCTTGAGG | |||
| FtGUX5-qf | TTGGAATGAAGCCGTGGGTT | |||
| FtGUX5-qr | ATCCATCTTCTCCGTCATCG | |||
| FtGXM1-qf | GCATAGGAATCTCCCGAAG | |||
| FtGXM1-qr | CATCACATCTGTCTCACCG | |||
| FtF5H-qf | TAGTTGAGTGGGCAATGGC | |||
| FtF5H-qr | TTGGAGGTAGGGCAAGTTG | |||
| FtCooAmot1-qf | GCCCTGCTTTGCCTGTTC | |||
| FtCooAmot1-qr | ATCATCCGCTGCGACCAC | |||
| FtTBL31-qf | GAGTAGGGAATAGCAACGAC | |||
| FtTBL31-qr | GATCTGCTCTTTCCTCCATG | |||
| FtTBL33-qf | CCAACATACTGGGCATCAAG | |||
| FtTBL33-qr | CCACTGCTTCTTATGGATGG | |||
| FtGXM3-qf | GACGAAATGCCTCAATCTCC | |||
| FtGXM3-qr | GAATGACCGCTATCTACACC | |||
| FtSEC2-qf | TCGTAACCTTCTCACCACCA | |||
| FtSEC2-qr | TACCCATACCTGCCTTCATC | |||
| FtHLK-qf | ATGTTGTGGGACGTGTGGTT | |||
| FtHLK-qr | AAGCGATCCTCTGGTGATAG | |||
| RolA-f | ATGGAATTAGCCGGACTAAACG | |||
| RolB-r | ATGGATCCCAAATTGCTATTCC | |||
| Ags-f | CGGAAATTGTGGCTCGTTGTGGAC | 内参基因 | ||
| Ags-r | AATCGTTCAGAGAGCGTCCGAAGTT | |||
| VirC-f | GGCTTCGCCAACCAATTTGGAGAT | |||
| VirC-r | TTTTGCTCCTTCAAGGGAGGTGCC | |||
| VirD-f | ATGTCGCAAGGACGTAAGCCCA | |||
| VirD-r | GGAGTCTTTCAGCATGGAGCAA | |||
| P35S-f | ATCTCCACTGACGTAAGGGA | |||
| T35S-r | TTGCGGACTCTAGCATGG | |||
注:下划线表示attB位点。 |
1.2 FtNAC16过表达载体的构建
利用Gateway技术构建FtNAC16过表达载体,该方法涉及BP和LR反应。根据FtNAC16的CDs序列及Gateway克隆系统要求的attB重组位点特征(载体pDONR221含互补的attP位点),设计含attB1/attB2位点的BP引物(表1)。以前期克隆得到的FtNAC16的CDs为模板,利用BP反应构建入门克隆并整合在FtNAC16基因两端后进行胶回收(具体操作参照天根试剂盒说明书)。使用Invitrogen试剂盒 Gateway BP Clonase Ⅱ Enzyme Mix(Thermo Fisher公司)进行入门载体构建,反应体系为:目的基因3 μL,pDONR221 vector 1 μL,Gateway BP Clonase Ⅱ Enzyme Mix 1 μL,ddH2O补足10 μL。25 ℃条件下连接3 h后加入蛋白酶K终止反应。通过热激法将重组产物转化到大肠杆菌感受态细胞中,并将转化菌液涂布在含Kan的LB平板上,37 ℃培养过夜。最后,挑选单一菌落进行PCR验证,确认含有目的片段的阳性菌株,送往生工生物工程(上海)股份有限公司测序。
将测序正确的入门载体pDONR221用于提取质粒。使用Invitrogen试剂盒 Gateway LR Clonase Ⅱ Enzyme Mix进行LR反应,完成过表达载体构建。LR反应体系如下:pDONR221-FtNAC16 100 ng,pK7WG2D 200 ng,LR enzyme 2 μL,ddH2O补足10 μL。反应条件为:25 ℃连接10 h,连接完成后取出PCR管加入1 μL蛋白酶K溶液,37 ℃孵育10 min后终止反应。经过LR反应,产物转化到大肠杆菌中,菌液涂板在含Spec的LB平板上,37 ℃倒置培养过夜,挑选单克隆进行PCR检测,并选择3个阳性菌株以P35S-f为引物进行测序。将测序正确的过表达载体pK7WG2D-FtNAC16提取质粒,并于-20 ℃冰箱保存备用。
1.3 过表达载体转化发根农杆菌Ar A4感受态
将含有100 μL发根农杆菌Ar A4感受态细胞的离心管在冰上解冻,随后加入200 ng的重组过表达质粒DNA并轻柔吸打混匀,将混合物依次进行冰浴(10 min)、液氮(5 min)、37 ℃水浴(5 min)。在无菌超净台中加入800 μL不含抗生素的TY液体培养基,置于振荡培养箱中25 ℃、200 r/min条件下培养2~3 h,然后以6 000 r/min离心1 min。在无菌超净台中弃掉800 μL上清液,吸打混匀剩余菌液,取20 μL涂布在含Kan和Spec抗生素的TY固体培养基上,28 ℃倒置培养2~3 d。挑选单克隆菌落进行菌落PCR检测,筛选阳性菌株。
1.4 农杆菌介导苦荞毛状根的遗传转化
挑选‘品苦荞1号’的饱满种子,使用灭菌水浸泡30 min后去壳。在无菌超净台上将种子依次用75%的酒精处理10 s和5%的NaClO溶液浸泡10 min进行消毒,再用灭菌水冲洗3~4次。用无菌吸水纸吸干表面水分,将种子接种至不含抗生素的MS固体培养基中生长。培养条件为25±2 ℃,光照16 h,黑暗8 h,光照强度2 500~3 000 lux。
农杆菌侵染及阳性苗筛选参考许亚春[27]的方法,将转基因工程菌与TY液体培养基混合,28 ℃、200 r/min条件下振荡培养6 h。当菌液吸光度OD600达到0.2~0.6时,离心收集菌体,并用含300 μmol/L AS的MS液体培养基悬浮至OD600为0.2。将预培养的苦荞外植体浸入菌悬液中10 min,吹干后转移至MS共培养基上,25±2 ℃条件下暗培养3 d。漂洗外植体,晾干后接种到含50 mg/L Cep抗生素的MS选择培养基中,25±2 ℃条件下进行16 h光照、8 h黑暗培养,光照强度为2 500~3 000 lux,连续培养2周。选择生长旺盛的转基因毛状根,切段后放入含50 mg/L Cep抗生素的MS液体培养基中悬浮培养,直至毛状根铺满锥形瓶底部。
1.5 FtNAC16转基因苦荞毛状根的鉴定
1.5.1 DNA水平鉴定
利用植物基因组DNA提取试剂盒提取FtNAC16转基因苦荞毛状根的DNA。检测DNA浓度,并通过1.2%琼脂糖凝胶电泳检测DNA纯度。以P35S-f和扩增目的基因下游引物FtNAC16-r1为扩增引物,DNA为模板,利用高保真KOD酶进行PCR扩增。PCR反应体系为:模板DNA 50 ng,0.3 μmol/L正反向引物各1 μL,dNTP 1 μL,MgSO4 0.4 μL,KOD-Plus-Neo 0.2 μL,10×buffer 1 μL,用灭菌双蒸水补足10 μL。PCR扩增程序为:预变性94 ℃ 3 min;94 ℃ 15 s,56 ℃ 30 s,68 ℃ 90 s,30个循环;68 ℃延伸10 min。PCR 产物4 ℃条件下保存。
根据发根农杆菌Ar A4所携带的Ri质粒,针对其转移区和毒性区的相关基因(RolAB、Ags、VirC和VirD)设计特异性引物RolAB-f/r、Ags-f/r、VirC-f/r和VirD-f/r(表1),以DNA为模板,分别以RolAB-f/r、Ags-f/r、VirC-f/r和VirD-f/r为引物进行PCR扩增。PCR反应体系为:模板DNA 50 ng,0.3 μmol/L正反向引物各1 μL,dNTP 1 μL,MgSO4 0.4 μL,KOD-Plus-Neo 0.2 μL,10×buffer 1 μL,用灭菌双蒸水补足10 μL。PCR扩增程序为:预变性94 ℃ 3 min;94 ℃ 15 s,56 ℃ 30 s,68 ℃ 90 s,30个循环;68 ℃延伸10 min。PCR 产物4 ℃条件下保存。
1.5.2 番红固绿石蜡切片观察分析
利用体视显微镜(Nikon, Japan)对野生型及转基因毛状根进行石蜡切片观察,石蜡切片的制作方法如下。
固定:将野生型(wild type,WT)和转基因毛状根放置于FAA固定液中,4 ℃固定24 h。
洗脱:以不同体积分数(70%、80%、90%、100%)的乙醇溶液进行逐步洗脱处理,每步1 h。
透明:在不同体积分数(25%、50%、75%、100%)的氯仿-乙醇溶液中进行透明处理,每步2~3 h。
浸蜡:分别在42 ℃、56 ℃、60 ℃的不同质量分数(50%、70%、100%)石蜡溶液中浸润1 h。
包埋:将材料放置于模具上,倒入60 ℃的石蜡溶液,置于冰上使石蜡完全凝固。
切片:使用自动切片机(LeicaRM2016)将包埋好的蜡块进行切片。
染色:进行番红固绿染色,利用荧光倒置显微镜(Olympus)观察毛状根内部结构。
1.5.3 扫描电镜观察分析
使用PBS对野生型及转基因毛状根表面进行清洗,将样本置于电镜固定液中室温固定2 h,以不同体积分数的乙醇溶液分级脱水,体积分数为30%、50%、70%、80%、90%、95%、100%的乙醇各脱水1次,每次15 min,再置于乙酸异戊酯中浸泡15 min。经过干燥处理后,将样品固定在导电碳膜双面胶上,并在离子溅射仪样品台上喷金30 s,最后利用S-3400N型扫描电子显微镜对样品进行观察和图像采集分析,利用Image-Pro Plus 6.0软件统计毛状根细胞壁的厚度。
1.6 FtNAC16转基因毛状根中次生壁组分含量分析
使用Van Soest法对野生型及转基因毛状根中纤维素、木质素和半纤维素的含量进行测定,具体操作参考段亚冰、徐苗均等的方法[28-29]。首先,制备中性洗涤剂、酸性洗涤剂和2.0 mol/L硫酸溶液,取1.0 g风干的毛状根,用中性洗涤剂和酸性洗涤剂进行处理,通过煮沸、抽滤、冲洗和烘干等步骤分离出中性洗涤纤维(neutral detergent fiber,NDF)和酸性洗涤纤维(acid detergent fiber,ADF)。随后将酸性洗涤纤维用72%硫酸消化,以测定酸性洗涤木质素(acid detergent lignin,ADL)和酸不溶灰分(acid-insoluble ash,AIA)。根据实验数据计算中性洗涤纤维、酸性洗涤纤维、半纤维素、酸性洗涤木质素和酸不溶灰分的含量。
1.7 FtNAC16在转基因苦荞毛状根中的表达分析
利用Omega植物总RNA试剂盒(广州飞扬生物工程有限公司)提取转基因苦荞毛状根根系总RNA。测定总RNA浓度,并通过1.2%琼脂糖凝胶电泳检测总RNA纯度。使用PrimeScriptTM Ⅱ 1st Strand cDNA Synthesis Kit试剂盒将获得的苦荞总RNA反转录,合成用于扩增基因全长CDs序列和实时荧光定量PCR(qRT-PCR)检测的第一链cDNA。提取野生型与转基因苦荞毛状根的总RNA,经过反转录RT-PCR获得cDNA。以不同转基因株系cDNA为模板,苦荞管家基因FtHLK为内参基因,测定并分析不同转基因株系中目的基因的相对表达量(以野生型为对照)。扩增程序为:95 ℃预变性 15 min;95 ℃变性 10 s,56 ℃退火20 s,72 ℃延伸30 s,40个循环;溶解曲线65 ℃ 0.05 s,95 ℃ 0.5 s。实验设置3个生物学重复和3个技术重复,用2-ΔΔCt方法计算相对表达量,具体参照孙小倩[30]的方法。
2 结果
2.1 FtNAC16过表达载体的构建
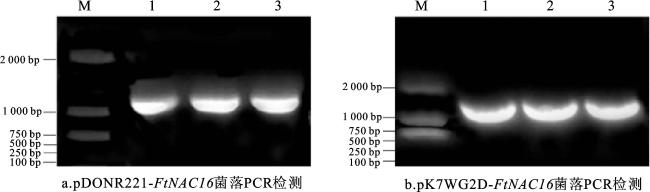
2.2 FtNAC16转基因毛状根阳性株系的鉴定
2.2.1 形态学鉴定
将重组质粒导入发根农杆菌Ar A4后,进行活化处理,并将其侵染转化生长7 d的苦荞幼苗。经3 d的暗培养后,幼苗被转移到正常光照条件下继续培养,约10 d后可见带有荧光的毛状根开始出现,挑选生长状况良好的毛状根切取2 cm转移到液体培养基中进一步培养为毛状根。毛状根的培养效果如图2a所示。
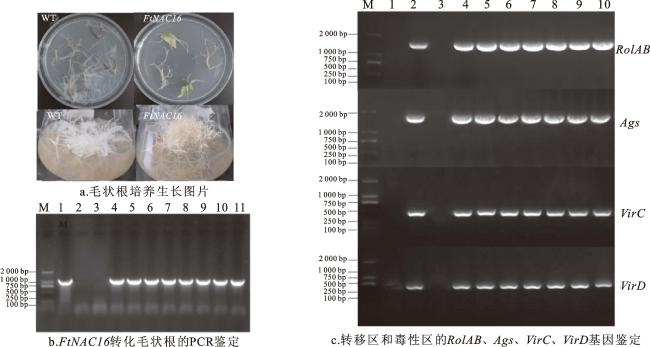
2.2.2 DNA水平鉴定
2.3 转基因毛状根中FtNAC16的表达分析
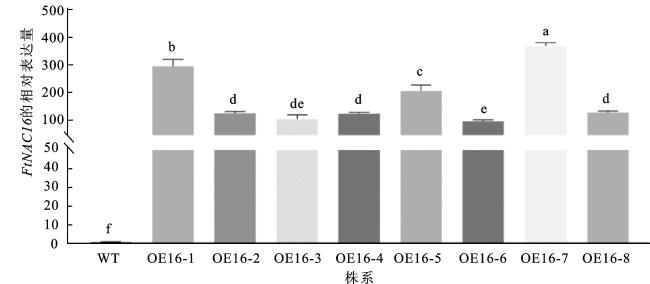
为获得FtNAC16高表达的转基因苦荞毛状根株系,以苦荞管家基因FtHLK为内参基因,分别对野生型和8个阳性株系进行qRT-PCR检测,结果如图3所示。FtNAC16在野生型毛状根中表达很低或不表达,而在转基因株系中均具有较高的表达。在FtNAC16转基因苦荞毛状根中选取表达量最高的3个株系OE16-1、OE16-5、OE16-7进行后续检测分析。
2.4 FtNAC16转基因毛状根的细胞学观察鉴定
2.4.1 番红固绿染色石蜡切片观察鉴定
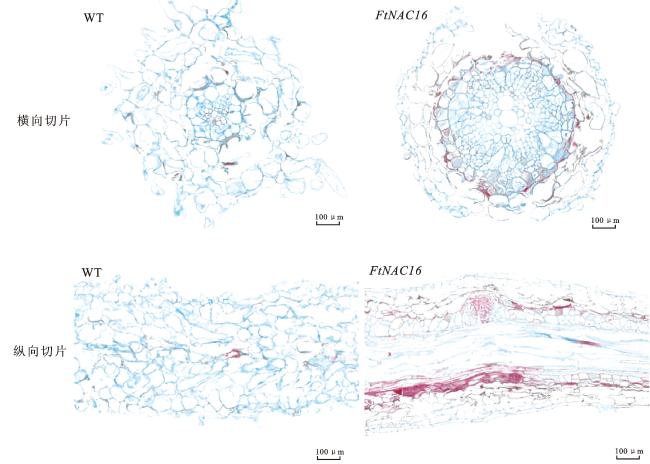
为探究FtNAC16转录因子在苦荞次生壁生物合成中的作用,选取转基因毛状根中表达量最高的株系OE16-7进行番红固绿染色石蜡切片观察,并以野生型毛状根作为对照。结果显示(图4),与野生型毛状根相比,FtNAC16转基因毛状根的红色区域更为显著,暗示FtNAC16过表达显著加速了细胞木质化和木栓化进程,进而促进了细胞壁中木质素和木栓质的积累。
2.4.2 扫描电镜观察鉴定

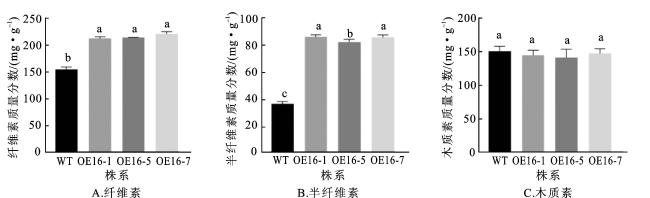
2.5 转基因毛状根中次生壁组分的含量分析
探究FtNAC16过表达对转基因毛状根次生壁组分含量的影响,选择表达量最高的3个转基因毛状根株系,并以野生型毛状根作为对照,对其纤维素、半纤维素和木质素含量进行测定。结果如图6所示,FtNAC16过表达显著增加了转基因毛状根中的纤维素和半纤维素含量,但对木质素含量无明显影响,表明FtNAC16过表达对次生壁中纤维素和半纤维素的生物合成具有促进作用。
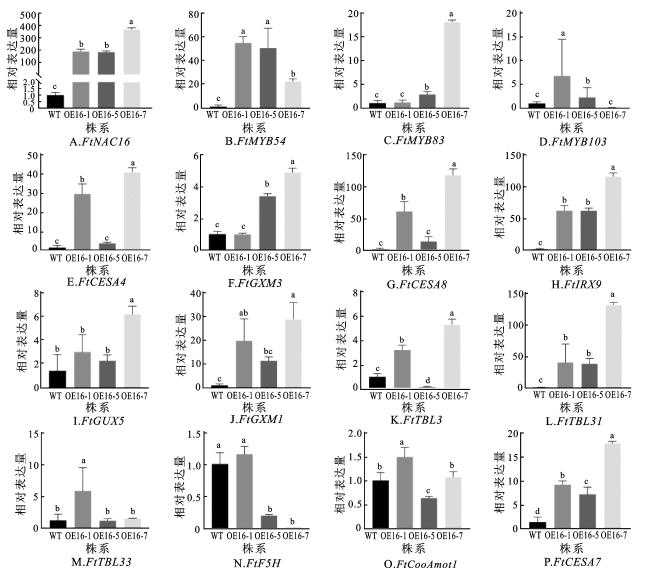
2.6 FtNAC16转基因毛状根次生壁生物合成途径调控下游基因的表达分析
进一步探究FtNAC16转录因子在苦荞次生壁生物合成中的作用机制,利用qRT-PCR分析野生型与FtNAC16过表达毛状根株系中相关基因的表达水平(图7)。以苦荞FtHLK为内参基因,对多个次生壁合成关键基因进行定量分析,包括FtMYB83、FtMYB103、FtMYB54等调控开关基因,FtCESA4、FtCESA7、FtCESA8等纤维素合成酶基因,FtIRX9、FtTBL3、FtGUX5、FtGXM1、FtTBL31、FtTBL33等半纤维素合成酶基因,以及木质素合成相关基因FtF5H、FtGXM3和FtCooAmot1。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
如图7所示,与野生型相比,二级调控结构基因FtMYB83和FtMYB54在FtNAC16过表达株系中显著高表达。此外,编码纤维素合成酶的关键结构基因FtCESA7和FtIRX9的表达水平也显著提升。在OE16-7转基因株系中,FtCESA7表达量为野生型的13.6倍,FtIRX9的表达量最高,是野生型的78.02倍(P<0.001),说明FtNAC16过表达显著促进了与纤维素合成相关下游基因的表达。以上结果证实FtNAC16通过激活次生壁合成调控网络,显著促进纤维素与半纤维素的生物合成,但其对木质素合成的调控具有复杂性。木质素合成相关基因(如FtF5H、FtCooAmot1)表达上调,部分转基因株系的木质素含量却下降,暗示木质素代谢受多基因协同调控。
3 讨论
大量研究表明,NAC转录因子(如拟南芥中的NST1、NST2、NST3/SND1)作为一级调控开关,在细胞次生壁的生物合成中扮演着关键角色[39] 。本课题组前期研究发现FtNAC16与拟南芥等植物中参与次生壁合成调控的NAC家族成员属于同一亚家族,即NST亚家族,并与杨树PtrWND1B和PtrWND2B、巨桉EgrWND1、玉米ZmNST3和ZmNST4等聚为一支[26]。在拟南芥中,NAC转录因子AtNST1、AtNST2和AtNST3通过调控次生壁合成相关的MYB转录因子(如MYB83、MYB103)和结构基因(如CESA4、CESA7、CESA8)的表达,促进次生壁加厚[40]。作为植物次生壁合成调控的关键NAC转录因子NST1、NST2和NST3/SND1的同源基因,FtNAC16可能在苦荞中发挥类似功能。
研究发现,拟南芥中由AtNST1、AtNST2和AtNST3/AtSND1调控的下游基因AtMYB83、AtMYB103、AtMYB54和纤维素合成结构基因AtCESA4、AtCESA7、AtCESA8在苦荞中的同源基因FtMYB83、FtMYB103、FtMYB54、FtCESA4、FtCESA7、FtCESA8在苦荞籽粒不同发育时期的表达模式与FtNAC16高度一致。这些结果暗示FtNAC16可能通过调节次生壁合成相关的下游调控基因和结构基因的表达,促进苦荞果壳次生壁加厚。
在其他植物中,NAC转录因子同样在次生细胞壁生长调控中扮演关键角色。在水稻中,OsSND2作为AtSND2的同源基因,对纤维素合成起正向调控作用。OsSND2在水稻节间和穗间区域高度表达,其过表达会导致叶片卷曲、纤维素含量上升以及与次生细胞壁相关的基因表达水平升高[41]。在棉花中,已鉴定出7个与次生细胞壁形成相关的NAC转录因子GhFSN1~7,其中GhFSN1作为正向调控因子,能够激活下游的次生细胞壁相关基因,进而参与调控棉花纤维的次生细胞壁形成[42];而GhFSN5作为负向调控因子,在拟南芥中异源表达后,会导致转基因植株的花序茎和根部纤维素和木质素含量下降[43]。在植物体内,NST1和NST3转录因子能够促进花粉内膜次生壁的增厚,也能促使下胚轴的次生木质部和次生壁增厚。这3个因子共同作用于下游SND2和SND3基因,促进木质素合成及次生壁增厚。
本研究对野生型和过表达FtNAC16的转基因毛状根进行qRT-PCR分析。结果显示,与野生型相比,FtNAC16基因在转基因毛状根株系中的表达水平显著上调。尽管qRT-PCR分析仅能揭示转录水平上的变化,表明过表达会导致该基因表达量的显著增加,但尚未明确该基因在次生壁合成过程中的具体作用机制。
为进一步探索FtNAC16的功能,本研究选择表达量最高的3个转基因株系OE16-1、OE16-5、OE16-7,检测其纤维素、木质素和半纤维素的含量,并将表达量最高的转基因株系OE16-7进行番红固绿石蜡切片和扫描电镜观察分析,同时测定其细胞壁厚度。检测结果显示,FtNAC16转基因毛状根的木质化和木栓化程度增强,纤维素、半纤维素含量均显著高于对照组,推测FtNAC16能够促进苦荞果壳中次生壁组分(纤维素和半纤维素)的合成及积累。通过对苦荞转基因毛状根进行功能验证,发现FtNAC16能够有效促进苦荞次生壁合成相关调控基因和结构基因的表达,但在部分转基因株系中,某些木质素合成相关基因的表达量显著上调,但木质素含量较野生型降低,推测木质素的生物合成过程可能受多个基因的复杂协同调控。
虽然本研究成功构建出FtNAC16基因的过表达载体,并通过苦荞毛状根转化对其进行了初步功能测试,但尚未在苦荞果壳中验证其功能。未来研究工作可聚焦于建立一套能够将苦荞愈伤组织诱导成完整植株的遗传转化体系,进而研究FtNAC16在苦荞果壳次生壁生物合成中的调控作用。
4 结论
本研究通过发根农杆菌介导的转基因毛状根体系,探究苦荞次生壁调控候选转录因子FtNAC16的功能。结果显示,过表达FtNAC16可以显著提高转基因毛状根细胞壁的厚度及纤维素、半纤维素的含量,并激活次生壁合成相关MYB转录因子及结构基因的表达,表明FtNAC16可能通过调控下游MYB因子及合成基因的表达,促进细胞壁增厚和组分积累。该研究为解析苦荞次生壁合成机制及果壳性状改良提供了理论基础。