

生物多样性是指在一定区域内所有生物资源的自然组合[1]。随着生物多样性丧失程度的不断加深,保护生物多样性已成为生态学研究的重点之一,深入研究生物多样性的形成和维持机制是生物多样性保护的重要方面[2]。喀斯特地区独特的生态系统和复杂的地形地貌形成了异质性极高的喀斯特生境,喀斯特生境中的生态因子受地形变化的控制,例如坡度、坡向、海拔等,进而维持着喀斯特地区群落植被的生物多样性[3-4]。
物种多样性是生物多样性最直观的表现形式,其能对一定区域内的物种资源丰富程度进行客观评价[5-6]。物种多样性是群落组成结构及抵抗外界干扰能力强弱的核心指标之一,可以很好地反映出群落的资源利用情况和对环境的适应能力,对生态系统的可持续发展具有重要意义[7-8]。生物多样性保护既要重视区域物种多样性的动态变化,又要掌握物种之间的进化信息,保护进化历史独特而又丰富的区域[9-10]。系统发育多样性可以提供物种之间的亲缘信息,并包含物种性状和生态位信息,能够用于推测历史发展和进化过程对群落的影响,进而能很好地解释生物多样性分布格局[11-13]。结合物种多样性和系统发育多样性可以有效解释群落在面对生境改变时的适应策略,为生物多样性保护提供了新视角[9,14]。
喀斯特地貌是可溶性岩石(主要为碳酸盐岩石)在侵蚀、溶蚀等地质作用下所形成的一系列特殊地貌。喀斯特地貌复杂多样,具有地势较高、坡度较陡、土壤物质和岩石易被淋溶、地下径流多、保水保肥能力差等特点,该地区植被生存条件极为严峻[15-16]。贵州省南部的茂兰国家级自然保护区具有同一纬度带上保存最为完整的非地带性喀斯特植被[17-18]。该保护区拥有错综复杂的地形,诸如山顶、山脊、山谷、鞍部、盆地、山坡等,为植物生存提供了丰富的小生境,使得茂兰国家级自然保护区具有极为复杂的生物多样性组合[19-20]。如今已有大量学者从不同角度对不同区域的物种多样性和系统发育多样性进行研究。基于山地生态系统的研究表明,气候变量的影响导致系统发育多样性与海拔呈负相关关系,在高海拔地区物种丰富度下降,系统发育多样性增加[21]。对高寒草甸生态系统进行研究发现,植被系统发育多样性随着海拔的增加整体呈下降趋势,且物种丰富度与系统发育多样性指数存在极显著正相关关系[22]。上述地形因子对植被系统发育多样性的研究主要集中于非喀斯特地貌地区[23],较为单一。喀斯特地区在大地形地貌尺度下,物种多样性更多受到小地形地貌及生境异质性的影响[24],对喀斯特地区不同地形之间的木本植物物种多样性及系统发育多样性研究较少。
基于此,本研究以贵州省茂兰国家级自然保护区的中国南方喀斯特森林生态系统茂兰动态监测样地(以下简称茂兰大样地)中不同地形的样方为研究地,选取山顶、山脊、山谷、鞍部、盆地、山坡6种地形,以木本植物为研究对象,从系统发育多样性和物种多样性两方面进行分析,以期明晰不同地形对植物群落物种多样性与系统发育多样性的影响,地形因子驱动植物群落物种多样性与系统发育多样性的空间分布特征,以及物种多样性与系统发育多样性指数之间的相关性,为喀斯特地区生态恢复以及生物多样性保护提供科学依据。
1 研究区概况
研究区位于茂兰国家级自然保护区(25°09'20″~25°20'50″ N,107°52'10″~108°45'40″ E),该保护区位于贵州省黔南布依族苗族自治州南部的荔波县,总面积约为20 000 hm2。区内海拔430~1 078.6 m,平均海拔800 m以上,为典型的喀斯特峰丛地貌,碳酸盐岩连片裸露,土层浅薄,土壤以黑色石灰土为主,pH 7.5~8.0,有机质和全氮含量丰富。该保护区以中亚热带季风温润气候为主,年均气温15.3 ℃,年均降水量1 750 mm,年均相对湿度83%。保护区内有喀斯特地貌区少有的原生性强、连片面积大且保存完整的喀斯特森林[25]。茂兰大样地位于茂兰保护区缓冲区与实验区交界处,地形极度复杂多变,岩石裸露率达90%以上,典型植被类型为非地带性常绿落叶阔叶混交林,植被群落比较成熟稳定,物种组成丰富,常见种有香叶树(Lindera communis)、齿叶黄皮(Clausena dunniana)、小果十大功劳(Mahonia bodinieri)等。
2 研究方法
2.1 样地设置
根据美国热带雨林研究中心(Center for Tropical Forest Science,CTFS)技术规范在茂兰大样地设置625个20 m×20 m的喀斯特森林植物群落样方,占地面积25 hm2(500 m×500 m)。根据喀斯特环境特点选择洼地、山谷、山坡、鞍部、山脊、山顶6个地形的样方进行调查(表1)[19],调查总面积为8 800 m2,调查时间为2022年6月至2023年3月。对样方内所有胸径大于等于1 cm的木本植物(包括木质藤本)每木调查,记录种名、高度、胸径等指标。植物种名参考物种2000(Species 2000 China Node,http:∥www.sp2000.org.cn/)。鞍部呈现马鞍形,是山脊上一个特殊部位,通常情况下,山脊延伸的尽头与鞍部相连,鞍部两侧向上延伸为山脊,以该特点来区分山脊和鞍部。山顶所在局部区域内呈现出局部地势高、陡峭程度大、相对高差大的特点,而山坡作为山脊、山谷等基本地貌间的连接部位,由坡度不断变化的倾斜面组成。在确定样方面积时,该样方和水平面的夹角所形成的投影面积达到20 m×20 m。
表1 不同地形的样方情况Tab.1 The quadrat situation of different terrains |
| 地形 | 样方数/个 | 样方面积/m2 | 物种数 | 密度/(个·m-2) |
|---|---|---|---|---|
| 洼地 | 4 | 400 | 102 | 0.434 |
| 山谷 | 3 | 400 | 115 | 0.598 |
| 山坡 | 4 | 400 | 126 | 0.721 |
| 鞍部 | 5 | 400 | 152 | 0.809 |
| 山脊 | 3 | 400 | 112 | 0.962 |
| 山顶 | 3 | 400 | 100 | 1.494 |
2.2 系统发育树构建
2.3 物种α多样性指数计算
采用Simpson优势度指数(Simpson dominance index,D)、Pielou均匀度指数(Pielou evenness index, J)、物种丰富度指数(species richness, S)衡量样方群落的物种多样性。物种多样性指数可以体现群落物种多样化水平,多方面反映该区域物种的丰富程度及群落稳定性,由R软件中的“picante”程序包计算完成。其中,木本植物的相对重要值=(相对频度+相对密度+相对胸高断面积)/3[24]。
$S=n。$
式中: n为样方内植物群落的总物种数量; Pi为第i个物种的相对重要值。
2.4 系统发育α多样性指数计算
采用Faith’s系统发育多样性指数(Faith’s phylogenetic diversity index,F)、平均成对系统发育距离(mean pair phylogenetic distance,MPD)、平均最近种间系统发育距离(mean nearest taxon phylogenetic distance,MNTD)衡量样方群落的系统发育多样性。Faith’s系统发育多样性指数被定义为群落内所有物种进化分支长度之和[29]。平均成对系统发育距离提供所有系统发育距离的平均值。平均最近种间系统发育距离仅包含物种与其亲缘关系最近物种的平均进化距离[30]。系统发育多样性指数由R软件中的“picante”程序包计算完成。
2.5 数据分析
对所有多样性数据进行正态分布检验和方差齐性检验,检验后符合正态分布和方差齐性。对各个地形间的物种多样性指数和系统发育多样性指数的差异水平采用单因素方差分析(one-way ANOVA)和最小显著差(least significant difference,LSD)进行多重比较分析。运用Pearson相关性分析物种多样性与系统发育多样性之间的关系。所有统计分析的显著水平设定为P<0.05。采用R软件的“ggplot2”“vegan”程序包对不同地形的多样性指数进行单因素方差分析[31]。在Chiplot(https:∥www.chiplot.online/)中对物种多样性指数和系统发育多样性指数进行相关性分析。使用RTK在样方四角定位,以插值得到1 m精度的数字高程模型(digital elevation model,DEM)数据为基础,在ArcGIS中依据数字高程模型使用Spatial Analyst Tools的表面分析获取各样方的坡度、坡位、曲率数据,同时使用RTK获取海拔数据。使用z-score法进行标准化作为地形因子,其中坡向为定类数据,没有进行标准化。在使用地形因子进行分析之前,为避免多重共线性的影响,使用Spearman相关性系数评估环境变量的冗余度,去除相关强度高的坡位(Spearman ρ2>0.7),保留坡度、坡向、曲率、海拔共4个地形因子。使用“rdacca.hp”程序包中的层次分割法评估地形因子的相对重要性以及它们对多样性指数的单独效应,“rdacca.hp”将层次分割法算法引入典范分析来获取一个典范分析解释变量的解释率,并实现不限解释变量数量限制的变差分解。层次分割法强调基于所有可能子集模型下各解释变量的单独效应,等同于解释变量的边际效应加上与其他解释变量的共同效应的平均分配值,以确定解释变量在所有子集模型中占主导地位的顺序[32]。
3 研究结果
3.1 不同地形的群落物种组成
如图1所示,在本研究已调查的所有样方内共出现有70科、154属、269种物种。其中,洼地102种、山谷115种、山坡126种、鞍部152种、山顶100种。
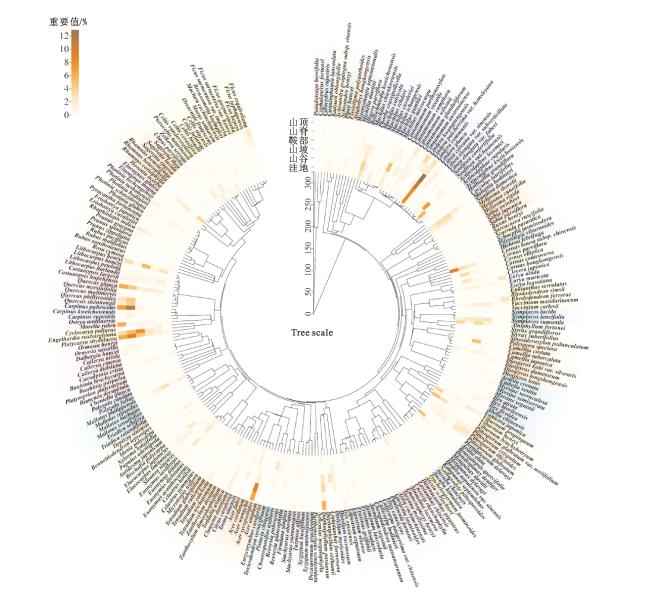
取各个地形中重要值最大的前3个物种表示该地形的优势种(表2),其中,山脊的优势种为云贵鹅耳枥(Carpinus pubescens)、化香树(Platycarya strobilacea)、角叶槭(Acer sycopseoides),鞍部的优势种为香叶树(Lindera communis)、黄杞(Engelhardia roxburghiana)、灰柯(Lithocarpus henryi),山顶的优势种为云贵鹅耳枥、化香树、角叶槭,洼地的优势种为小花梾木(Cornus parviflora)、香叶树、柳叶黄肉楠(Actinodaphne lecomtei),山谷的优势种为香叶树、四川新木姜子(Neolitsea sutchuanensis)、黔桂润楠(Machilus chienkweiensis),山坡的优势种为香叶树、小果润楠(Machilus microcarpa)、突肋茶(Camellia costata)。可以发现,香叶树和化香树在6个地形中都占有较大比例,这是因为喀斯特地区土层瘠薄,保水能力差,香叶树和化香树耐旱耐浅土,生态位宽度较宽,能够很好地适应喀斯特地区环境。
表2 不同地形中重要值排名前5的物种Tab.2 The top five species in terms of importance value in different terrains |
| 地形 | 物种 | 重要值/% |
|---|---|---|
| 山脊 | 云贵鹅耳枥(Carpinus pubescens) | 11.42 |
| 化香树(Platycarya strobilacea) | 9.19 | |
| 角叶槭(Acer sycopseoides) | 9.10 | |
| 黄杞(Engelhardia roxburghiana) | 5.17 | |
| 青冈(Quercus glauca) | 4.60 | |
| 鞍部 | 香叶树(Lindera communis) | 12.92 |
| 黄杞(Engelhardia roxburghiana) | 7.89 | |
| 灰柯(Lithocarpus henryi) | 5.42 | |
| 化香树(Platycarya strobilacea) | 4.24 | |
| 脉叶虎皮楠(Daphniphyllum paxianum) | 3.64 | |
| 山顶 | 云贵鹅耳枥(Carpinus pubescens) | 7.04 |
| 化香树(Platycarya strobilacea) | 6.35 | |
| 角叶槭(Acer sycopseoides) | 6.31 | |
| 青冈(Quercus glauca) | 6.29 | |
| 杨梅(Morella rubra) | 6.14 | |
| 洼地 | 小花梾木(Cornus parviflora) | 9.21 |
| 香叶树(Lindera communis) | 8.48 | |
| 柳叶黄肉楠(Actinodaphne lecomtei) | 5.66 | |
| 短序荚蒾(Viburnum brachybotryum) | 5.11 | |
| 栎叶罗伞(Brassaiopsis quercifolia) | 4.78 | |
| 山谷 | 香叶树(Lindera communis) | 8.29 |
| 四川新木姜子(Neolitsea sutchuanensis) | 7.47 | |
| 黔桂润楠(Machilus chienkweiensis) | 4.10 | |
| 灯台树(Cornus controversa) | 3.32 | |
| 小花梾木(Cornus parviflora) | 3.22 | |
| 山坡 | 香叶树(Lindera communis) | 10.93 |
| 小果润楠(Machilus microcarpa) | 5.34 | |
| 突肋茶(Camellia costata) | 5.16 | |
| 赤杨叶(Alniphyllum fortunei) | 3.48 | |
| 小果十大功劳(Mahonia bodinieri) | 3.37 |
3.2 物种多样性与系统发育多样性的变化
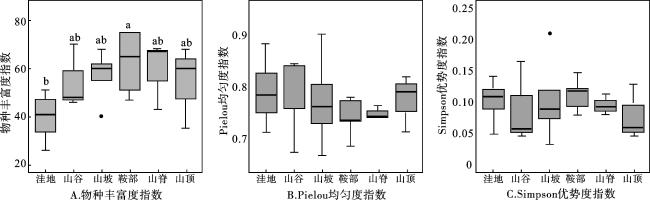
由图2可知,在6种地形木本植物群落中,鞍部的物种丰富度指数最大,洼地的物种丰富度指数最小。最小显著差分析结果表明,Pielou均匀度指数、Simpson优势度指数在不同地形之间无显著差异,但物种丰富度指数在洼地与鞍部之间存在显著差异(P<0.05)。
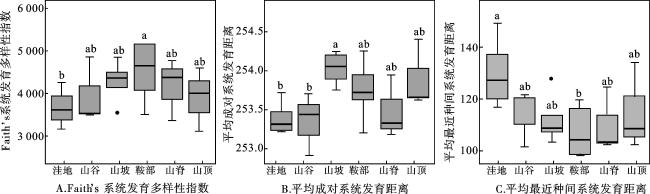
物种数越多,Faith’s系统发育多样性指数越大。鞍部的Faith’s系统发育多样性指数最大,洼地最小,该指数随地形变化的趋势与物种丰富度指数基本一致。山坡的平均成对系统发育距离最大;洼地的平均最近种间系统发育距离最大(图3)。最小显著差分析结果表明,Faith’s系统发育多样性指数、平均成对系统发育距离、平均最近种间系统发育距离在不同地形之间存在显著差异,平均成对系统发育距离在山坡与洼地、山谷之间差异显著,平均最近种间系统发育距离在洼地与鞍部之间差异显著。
3.3 地形因子对物种多样性和系统发育多样性变化的影响
由图4可以看出,4个地形因子的相对重要性排序依次为坡向(36.24%)、坡度(7.18%)、曲率(-3.96%)、海拔(-4.46%)。坡向、坡度、曲率、海拔的边际效应分别为45.87%、14.32%、-3.42%、-3.77%。坡向与坡度的共同效应最低,为-15.23%;坡度与曲率的共同效应最高,为1.52%。坡向、坡度、海拔三者的共同效应最高,为1.21%;坡向、坡度、曲率三者的共同效应最低,为-0.93%。坡向与坡度的共同效应出现负值,形成了坡向的边际效应(45.87%)大于单独效应(36.24%),坡度的边际效应(14.32%)大于单独效应(7.18%)。四者共同解释了多样性指数总变差的35%(校正R2=0.35)。4个地形因子对多样性指数的影响均不显著。
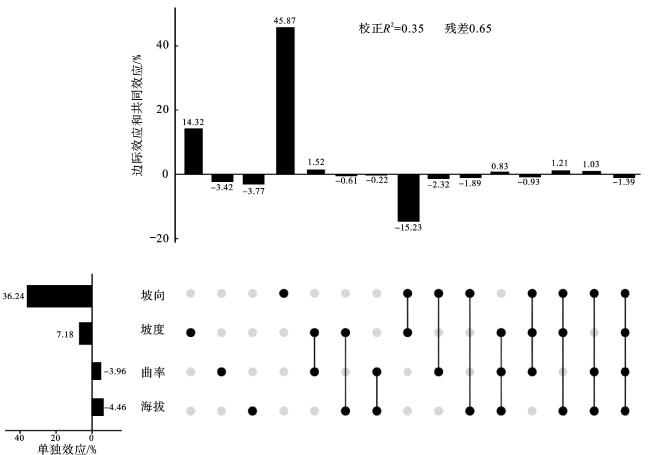
图4 4种地形因子对样方内木本植物群落物种多样性指数和系统发育多样性指数影响的相对重要性注:变差分解和层次分割分析结果使用UpSet图呈现。右侧点阵图中,每行对应一个地形因子。每列孤立黑点表示各地形因子的边际效应,多点间连线表示地形因子间的共同效应,各组分解释的变差百分比(来自变差分解)展示在上方柱形图中。左侧柱形图为各地形因子的单独效应(来自层次分割),其值等同于该地形因子的边际效应加上与其他地形因子的共同效应的平均分配值。 Fig.4 The relative importance of four topographic factors on species diversity and phylogenetic diversity of woody plant communities in the plots |
3.4 物种多样性和系统发育多样性的相关性
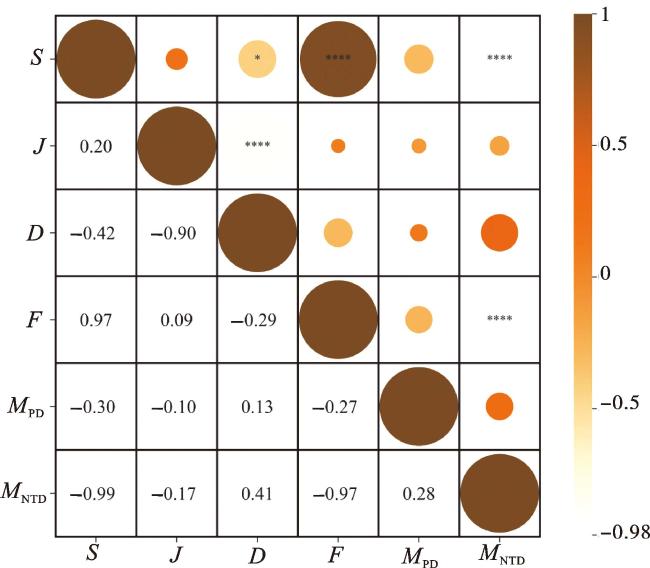
由图5可知,物种丰富度指数与Faith’s系统发育多样性指数极显著正相关,与平均最近种间系统发育距离极显著负相关,与Simpson优势度指数显著负相关。Pielou均匀度指数与Simpson优势度指数极显著负相关,Faith’s系统发育多样性指数与平均最近种间系统发育距离之间极显著负相关。平均成对系统发育距离与物种多样性指数之间相关性不显著。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
4 讨论
系统发育多样性能够在一定程度上预测群落的动态变化过程,对揭示群落生态变化有重要意义[35]。本研究中,Faith’s系统发育多样性指数与物种丰富度指数正相关,Faith’s系统发育多样性指数越大,表明群落中共同祖先越多,进化历史差异性就越大,生物多样性就会更加丰富[29]。平均成对系统发育距离与平均最近种间系统发育距离在不同地形中表现不同。从洼地到山谷再到山坡的平均成对系统发育距离表现出增大趋势,说明这三处地形样方内物种之间的系统发育距离逐渐增大,亲缘关系也越来越远;而从山坡到鞍部至山脊的平均成对系统发育距离表现出减小趋势,说明这三处生境样方内物种之间的系统发育距离逐渐减小,亲缘关系也越来越近。在所选地形中平均成对系统发育距离整体呈现不均匀分布格局,表明各地形树种的平均系统发育距离是不相同的,在历史进程中各树种受到的干扰程度不同,各地形中的群落进化不同步。山坡的平均成对系统发育距离最大,在生境异质性的影响下,来自山坡的古老物种在鞍部、山脊生境的繁殖和生存会变得更加艰难。高资源生产力有利于古老群落生存[36],且有研究指出,在喀斯特地区地形因子是造成土壤养分空间变异的重要因素[37],推测在喀斯特地貌中山坡生境类型具有较高的资源生产力,使其具有更加稳定的进化历史,为木本植物群落提供了生态保障[38]。洼地接受太阳辐射较少,局部地段会出现季节性积水,导致洼地中所生长和繁殖的大多数植被为耐阴性较强的植被,这可能是比较极端的环境对植物进化距离起到了一定的抑制作用,使洼地的物种分化时间较短。地形作为喀斯特地区极为重要的生态因子,通过影响光照、降水、土壤养分等的分配情况,形成了植被生长发育所需资源的差异,为在不同地形的喀斯特生境植被恢复中选择适宜树种提供了很好的参考。
坡向小尺度范围内的变化影响地表植被对光照时长、光照强度的接受程度,导致土壤水热因子发生变化[20,39]。本研究中,坡向的相对重要性最大,起主导作用,海拔的相对重要性最小,而在大多数研究中,海拔对群落α多样性有直接影响或占主导作用[40],推测造成这种结果的原因是:本研究所选样方地处喀斯特地区独特的峰丛地貌,相对高差不大,而大部分样方之间生境异质性强,这在很大程度上导致坡向多样化,使光照时长和强度产生差异。坡向和坡度二者的边际效应明显大于其单独效应,且二者的共同效应为负值,推测坡度影响地表的降水滞留量以及对土壤养分的淋溶,坡向影响光照时间与强度,二者相互作用,共同影响物种α多样性格局。所选4个地形因子对多样性指数的影响都不显著,这可能是由于喀斯特地区地形多样且高度破碎化,并伴随着诸多不稳定因素(气候因子、土壤理化性质等),群落内部存在极为复杂的相互联系。本研究仅探索了地形因子的影响,并没有涉及到土壤养分、温度、降水等环境因子的影响,今后研究需要补充更加丰富的地形因子以及环境因子对喀斯特地区多样性指数的影响。
本研究中,Faith’s系统发育多样性指数与物种丰富度指数正相关。已有大量研究证实,系统发育多样性与物种多样性之间存在相关性,如喀斯特地区乔木层、灌木层、草本层群落植物的Faith’s系统发育多样性指数与物种丰富度指数、Simpson优势度指数之间高度相关[3]。这种相关性在进化树不平衡或者存在独特物种分布狭域的情况下会降低,如在南非好望角森林群落研究中,发现Faith’s系统发育多样性指数与物种丰富度指数所对应的物种多样性区域不匹配,这与进化树不平衡有关[41]。本研究中,Faith’s系统发育多样性指数和Simpson优势度指数、Pielou均匀度指数的相关性不显著,推测这是由于该喀斯特地区生境异质性极高,不同树种的分布范围差异较大所致。在大部分研究中,随着Pielou均匀度指数的增加,优势种在群落中的竞争力降低,导致对环境资源的垄断减少,从而提供出更多生态位,其他物种会迅速占据,增加了群落的种间系统发育距离,导致平均成对系统发育距离增加[11,22]。本研究中平均成对系统发育距离与物种多样性指数之间无明显相关性,Pielou均匀度指数变化平缓,平均成对系统发育距离在各地形之间呈现先上升后下降再上升的变化趋势。这可能是喀斯特地区地形复杂,形成了多样化的小生境,植被为适应不同小生境而出现生态位分化所导致的。在该喀斯特地区物种多样性与系统发育多样性之间相关性不高,在对喀斯特地区进行生态恢复和生物多样性保护过程中,需要根据重点保护情况,对物种多样性和系统发育多样性指数进行综合考虑和合理选择。
5 结论
通过分析喀斯特森林生态系统茂兰动态监测样地中不同地形的物种多样性和系统发育多样性,发现在茂兰大样地中,基于不同地形的物种多样性指数差异不显著,只有物种丰富度指数在洼地与鞍部之间存在显著差异,Simpson优势度指数与Pielou均匀度指数均不显著。不同地形之间的系统发育多样性指数存在显著差异,Faith’s系统发育多样性指数和平均最近种间系统发育距离均在洼地与鞍部之间存在显著差异,平均成对系统发育距离在山坡和洼地、山谷之间存在显著差异。多样性指数受坡向、坡度为主的地形因子的影响,坡向是影响多样性指数的主导因子,坡度其次,4类地形因子对多样性指数均无显著影响。样地中的物种多样性指数与系统发育多样性指数整体相关性不明显。