

2019年末全球遭遇了前所未有的公共卫生挑战:由严重急性呼吸综合征冠状病毒2(SARS-CoV-2)引发的新型冠状病毒肺炎(COVID-19)疫情迅速席卷全球,不仅对人类健康造成了巨大威胁,也对全球经济和社会结构产生了深远影响[1-2]。SARS-CoV-2作为冠状病毒家族的一员,其独特的生物学特性和复杂的复制机制成为研究热点。冠状病毒是一类具有包膜的单股正链RNA病毒,广泛存在于自然界中,能够感染包括人类在内的多种哺乳动物和鸟类[3]。这类病毒因其表面特征性的冠状突起而得名,其基因组结构复杂,编码多种蛋白质,这些蛋白质在病毒的复制、组装和致病过程中发挥着关键作用[4]。其中,nsps作为病毒复制酶-转录酶复合体的重要组成部分,对病毒生命周期至关重要[5-6]。
在SARS-CoV-2的nsps中,nsp9蛋白因其独特的生物学功能和在病毒复制过程中的核心地位而备受关注。nsp9是一种单链核酸结合蛋白,具有高度的保守性和特异性,能够通过与病毒RNA相互作用,参与病毒的复制和转录过程[7]。nsp9的缺失或功能异常往往会导致病毒复制效率显著下降,从而影响病毒的致病性和传播能力[8-9]。近年来,随着对SARS-CoV-2研究的不断深入,研究人员对nsp9蛋白的结构、功能和作用机制有了更加全面的认识。研究表明,nsp9在病毒复制酶-转录酶复合体中扮演着“桥梁”角色,能够通过与nsp8等其他nsps的相互作用,形成稳定的复合体结构,为病毒RNA合成提供必要的平台和环境[10]。此外,nsp9还能够与宿主细胞的某些蛋白质相互作用,如在SARS-CoV-2入侵宿主后,宿主SND1蛋白可募集nsp9蛋白进而促进新生病毒RNA的合成并充当蛋白质引物,为病毒的复制和致病创造有利条件[11]。除此之外,nsp9也可靶向TBK1蛋白并促进细胞因子的生成,在病毒感染模型中,nsp9诱导的细胞因子风暴加剧了小鼠的组织损伤和死亡[12]。鉴于nsp9在SARS-CoV-2复制和致病过程中的重要作用,针对该蛋白的研究不仅有助于揭示SARS-CoV-2的复制机制和致病机理,也可以为开发新型抗病毒药物和疫苗提供重要靶点和思路。
综上,对SARS-CoV-2中nsp9蛋白的研究具有重要科学意义和应用价值。随着研究技术的不断进步和研究内容的不断深入,有望更加全面地揭示nsp9蛋白的生物学特性和作用机制,为应对疫情挑战提供更加有力的支撑。因此,本研究综合利用多种实验方法,对nsp9蛋白的细胞表达定位及潜在的宿主互作蛋白进行分析,探究其对宿主细胞增殖和凋亡的影响,为进一步明确nsp9蛋白在SARS-CoV-2复制过程中的作用提供一定的实验基础。
1 材料与方法
1.1 主要材料
Hela及Hpaepic细胞系,购自商城北纳创联生物科技有限公司;HEK293T细胞为实验室自存。引物合成均在浙江有康生物科技有限公司完成。I-5 High-Fidelity PCR酶及感受态细胞,购自北京擎科生物科技股份有限公司。核酸及蛋白marker、细胞总RNA提取试剂、反转录试剂盒、Dpn1限制性内切酶,购自宝日医生物技术(北京)有限公司。质粒提取试剂盒、胶回收试剂盒、氨苄青霉素、考马斯亮蓝G250/R250、IPTG,购自生工生物工程(上海)股份有限公司。亲和层析介质GST及Ni,购自美国Cytiva(思拓凡)公司。Lipo3000转染试剂、IP裂解液,购自美国赛默飞世尔科技公司。CCK8细胞活力检测试剂、Anti-Flag磁珠,购自美国MedChemexpress生物科技公司。EdU增殖检测试剂盒、TUNEL检测试剂盒,购自南京诺唯赞生物科技股份有限公司。EGFP、Flag抗体,购自杭州华安生物技术有限公司。
1.2 主要方法
1.2.1 载体构建
表1 本研究所用引物信息Tab.1 Primer information used in the experiment |
| 引物名称 | 引物序列(5'-3') |
|---|---|
| TRAF2-F | CGACCGTTGGGGCTTTGTTC |
| TRAF2-R | CCCTGCCCACTTGTACAGTGAGAC |
| YWHAB-F | GAGTCAGGGAATGACAATGGA |
| YWHAB-R | CACAAAGCACGAGAAACATTAG |
| TRAF2-overlap-F | CTGGTGCCGCGCGGCAGCATGGCTGCAGCTAGCGTGA |
| TRAF2-overlap-R | GATGATGATTCCTCCTCCTCAGAGCCCTGTCAGGTCCAC |
| YWHAB-overlap-F | CTGGTGCCGCGCGGCAGCATGACAATGGATAAAAGTG |
| YWHAB-overlap-R | GATGATGATTCCTCCTCCTTAGTTCTCTCCCTCCCCA |
| pCDNA3.1-EGFP-F | GTTGTTTACGGGTATTTTATAAAAAATACGAGG |
| pCDNA3.1-EGFP-R | TATAAAATACCCGTAAACAACTTCAACCTTATCAC |
| nsp9-EGFP-F | GGAGGAGGAATCATCATCATGAACAACGAACTGAGCCCA |
| nsp9-EGFP-R | CTCCTCGCCCTTGCTCACCTGCAGACGAACGGTTGC |
注:表中粗体为该基因克隆引物,gene-overlap-F/R为基因亚克隆引物。 |
1.2.2 蛋白原核表达纯化
根据GeneBank所提供的序列进行目的基因克隆,并将其构建至pGEX-6p-1载体,载体转化至BL21(DE3)感受态细胞中进行蛋白诱导表达,转化方法参考感受态细胞说明书。
诱导方法如下:将过夜培养的5 mL菌液加入到含有抗生素的1 L LB液体培养基中,37 ℃、200 r/min条件下培养约4 h(菌液OD600值达到0.6~0.8),加入终物质的量浓度为0.5 mmol/L的IPTG,在16 ℃、200 r/min条件下继续培养18 h。在4 000 r/min、4 ℃条件下离心10 min,收集菌体,用裂解缓冲液重悬菌体后,使用高压均质机破碎至溶液稍透明且不黏稠。在15 000 r/min、4 ℃条件下离心 40 min,收集上清液进行GST或Ni亲和层析。经SDS-PAGE凝胶检测分析后再进行离子交换或凝胶过滤层析,获得纯度和性质较好的目的蛋白。
1.2.3 Pull-down实验
取GST或Ni珠子20 μL,用含50 mmol/L Tris、150 mmol/L NaCl、0.05% Tween 20的缓冲液轻柔地洗3次,随后加入纯化后的目的蛋白和nsp9(n目的蛋白∶nnsp9=1∶1),4 ℃条件下摇晃孵育30 min。在3 000 r/min、4 ℃条件下离心5 min,小心移去上清液,并用缓冲液充分洗涤5次。洗涤完成后除去上清液,加入含有2% SDS的1×loading buffer 20 μL,95 ℃煮10 min,进行SDS-PAGE凝胶电泳分析。
1.2.4 免疫共沉淀质谱(immunocoprecipitation mass spectrometry,IP-MS)
将生长状态良好的HEK293T细胞铺板至6 cm皿中,待汇合度达到80%时分别转染2.5 μg的对照质粒(pcDNA3.1)和pcDNA3.1-Flag-nsp9质粒,48 h后弃去培养基,PBS清洗3次,继续加入免疫共沉淀裂解液,用细胞刮刀收集细胞并在冰上充分裂解30 min。12 000 r/min、4 ℃条件下离心15 min,吸取上清液进行BCA定量。取2 mg总蛋白进行免疫共沉淀实验,操作方法按照说明书中的要求。取上述免疫共沉淀后的蛋白样品5 μL,先进行银染鉴定,显色后进行观察分析,然后将剩余所有样品用于SDS-PAGE凝胶电泳,待蛋白样迁移至分离胶0.5 cm时,停止电泳,将胶取出用ddH2O清洗3遍,放入干净的器皿中进行样品切割,对凝胶样品进行酶解脱盐等处理后送至四川大学进行质谱检测。
采用Q Exactive组合型四极杆质谱仪结合EASY-nLC 1000液相色谱系统进行LC-MS/MS分析。使用C18色谱柱(120 mm×75 μm,5 μm)进行高效液相色谱,以70 000(AGC 3e6)分辨率获取全扫描质谱(350~1 800 m/z)。前20个多电荷离子通过高能量碰撞解离进行碎片化,子离子在分辨率为17 500(AGC 1e6)的轨道上测量。质谱条件如下:喷雾电压2 kV,加热毛细管温度250 ℃,标准化解离碰撞能量27%。全扫描和二级扫描的最大填充时间分别设置为20 ms和64 ms,动态排除时间设置为30 s。
细胞内免疫共沉淀验证的细胞培养处理方法同上,质粒转染为nsp9与互作蛋白同比例共转染,后续经蛋白质印迹(western blot,WB)鉴定。
1.2.5 细胞定位
第1天,将生长状态良好的HeLa进行细胞消化计数,铺12孔板;第2天,观察细胞密度及均匀度,待细胞密度达到80%时进行质粒转染,转染量为1 μg;第3天,将细胞消化下来转移至激光共聚焦小皿中进行培养;第4天,在荧光显微镜下观察细胞内绿色荧光蛋白的表达情况。
细胞核染色:配制Hoechst 33258染色液(质量浓度为0.5 μg/mL,PBS配制),弃去细胞培养基,PBS漂洗2次,加入足以覆盖细胞的Hoechst 33258染色液,常温孵育约15 min,弃去染色液,PBS清洗3次。利用激光共聚焦进行成像分析。
1.2.6 细胞增殖和凋亡检测
CCK8细胞活力检测、EdU细胞增殖实验及TUNEL细胞凋亡实验的操作方法均参考相应的试剂盒说明书。
2 结果与分析
2.1 SARS-CoV-2 nsp9蛋白的细胞定位
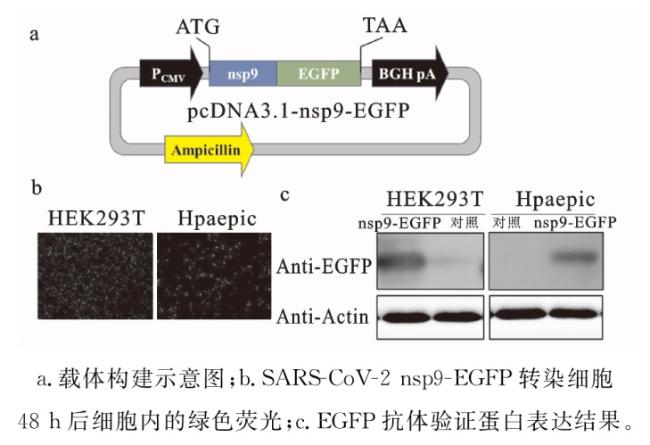
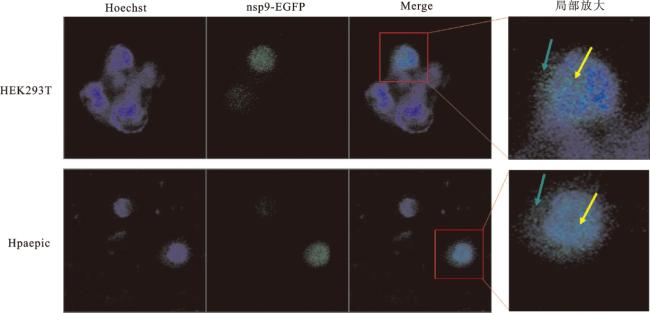
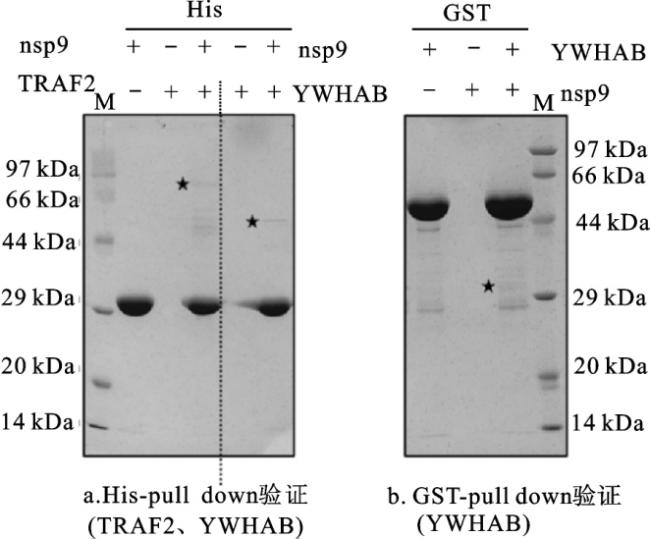
2.2 SARS-CoV-2 nsp9与宿主细胞蛋白相互作用
如图3所示,为明确SARS-CoV-2 nsp9在细胞内表达后是否会与宿主细胞蛋白发生相互作用, 在细胞中转染nsp9过表达质粒,利用免疫共沉淀质谱
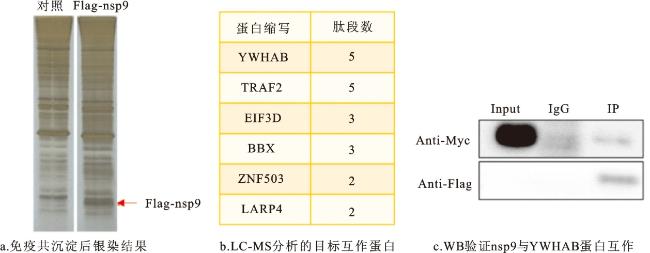
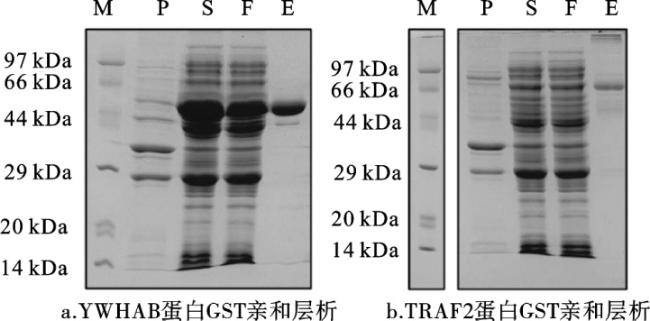
图4 与nsp9互作宿主细胞蛋白在大肠杆菌中的表达注:M代表marker;P代表沉淀;S代表上清;F代表穿透;E代表洗脱。 Fig.4 Expression of host cell proteins interacting with nsp9 in E. Coli |
2.3 SARS-CoV-2 nsp9对细胞增殖和凋亡的影响
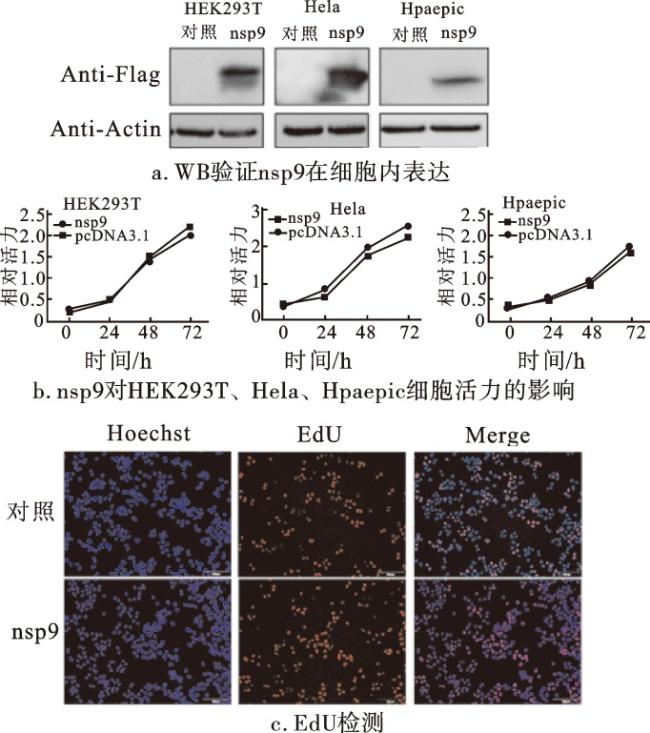
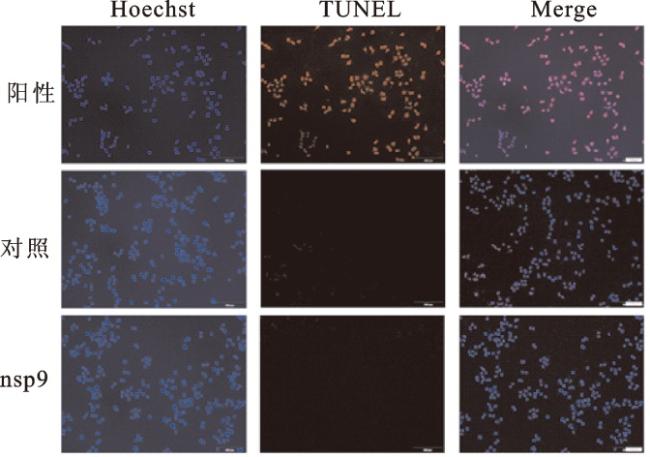
YWHAB蛋白(14-3-3 β蛋白)是14-3-3蛋白家族中表达量较高的成员之一,其参与细胞内多种信号通路和胞内蛋白跨膜转运等过程,也可以调节细胞增殖[14]。TRAF2蛋白是TRAF家族中较为经典的成员之一,在生物体免疫过程中起重要作用,并在细胞凋亡以及应激反应中起调节作用;其在多种组织细胞中均有表达,表明TRAF2蛋白在正常或病理过程中都可能发挥作用[15]。nsp9与YWHAB和TRAF2蛋白的互作提示其有可能参与细胞的增殖或凋亡过程。因此,本研究在细胞内过表达nsp9蛋白,并利用CCK8细胞活力检测和EdU细胞增殖实验验证nsp9对细胞增殖的影响,利用TUNEL细胞凋亡实验验证nsp9对细胞凋亡的影响。
上述实验在HEK293T、Hela和Hpaepic 3种细胞中进行,分别将nsp9转染以上3种细胞,并利用蛋白质印迹实验验证,结果证实nsp9在细胞内可以成功过表达(图6a)。采用CCK8细胞活力检测测定对照组和nsp9过表达质粒的HEK293T、HeLa、Hpaepic细胞增殖速率。由增殖曲线可以看出,nsp9对3种细胞的正常增殖均没有较大影响(图6b)。EdU染色结果进一步证实,nsp9过表达对细胞增殖无显著影响(图6c)。利用TUNEL细胞凋亡实验检测nsp9对细胞凋亡的影响,发现nsp9过表达后的细胞凋亡数量极少,与对照组相比无显著差异(图7)。以上结果提示,新冠病毒入侵细胞后,nsp9蛋白并不会显著影响细胞的增殖和凋亡。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 讨论
冠状病毒作为一类广泛存在于自然界中的病毒家族,具有独特的生物学特性,其蛋白与宿主细胞蛋白之间的复杂互作,不仅决定了病毒的感染效率,还深刻影响着宿主的免疫反应和细胞功能。例如,SARS-CoV和SARS-CoV-2病毒的入侵开始于病毒表面刺突蛋白识别并结合宿主细胞上的血管紧张素转化酶2(ACE2)受体[16-17];病毒进入细胞后,其RNA基因组在宿主细胞因子的协助下进行复制和转录,同时宿主细胞会启动一系列免疫反应以清除病毒。然而,病毒通过其蛋白与宿主细胞蛋白的互作,可以抑制或逃避宿主免疫反应,如nsp1可以抑制宿主细胞的翻译过程,从而逃避宿主的免疫监视[18];ORF6与病毒nsp8蛋白互作从而参与病毒的复制并抑制干扰素生成[19]。
本研究通过IP-MS发现SARS-CoV-2 nsp9蛋白与细胞内YWHAB、TRAF2、BBX等多个蛋白可能存在相互作用,其中BBX、ZNF503定位在细胞核,nsp9可能在细胞核内与之相互作用并影响细胞进程,但本研究未能成功在细胞以及体外表达上述2个蛋白,从而未能充分证明nsp9与其发生互作。YWHAB和TRAF2蛋白为本研究证实的与nsp9存在一定互作的蛋白。其中,YWHAB为多功能蛋白,在细胞内可以与多种蛋白相互作用,参与细胞周期调控、细胞凋亡、细胞信号转导以及代谢调控等多种细胞生物学过程[20]。另一互作蛋白TRAF2是TRAF家族成员,在生物体的免疫过程中发挥重要作用,并对细胞凋亡以及应激反应具有调节作用[15]。研究表明,YWHAB和TRAF2蛋白对于病毒的入侵及复制具有重要作用。YWHAB能够与2型猪圆环病毒(PCV2)的ORF5蛋白发生相互作用,抑制ORF5和Cap基因的转录以及Cap蛋白的表达,最终抑制PCV2在细胞中复制[21]。在针对痘苗病毒[22]和牛病毒性腹泻病毒[23]的研究中,TRAF2能够促进病毒快速进入宿主细胞,并通过siRNA干扰其表达,致使细胞内病毒滴度显著下降。本研究通过多种实验手段发现,nsp9与YWHAB、TRAF2蛋白的互作并不能影响宿主细胞的增殖和凋亡,但受限于实验条件,nsp9与YWHAB、TRAF2的相互作用是否会影响SARS-CoV-2的入侵以及复制有待深入研究。
4 结论
本研究发现新冠非结构蛋白nsp9主要定位于细胞核,少量在细胞质;明确了nsp9与宿主YWHAB、TRAF2等蛋白存在潜在互作关系;并检测出nsp9未对细胞正常增殖及凋亡产生显著影响。研究结果为进一步探究nsp9和宿主蛋白的相互作用、明确nsp9在SARS-CoV-2复制过程中的具体作用机制提供了一定的实验基础。