

生物和非生物胁迫对作物的生长发育具有负面影响。非生物胁迫包括渗透胁迫(如干旱和盐胁迫)、高温和低温胁迫、重金属胁迫以及养分缺乏等,严重影响作物产量[1-2]。盐胁迫是影响植物生长发育的重要障碍,土壤中盐分过量会引发植物的渗透胁迫、离子中毒、失水、萎蔫、代谢紊乱以及膜系统受损等问题,严重时甚至会导致植物死亡[3-5]。
钙离子(Ca2+)作为植物体内的第二信使,是各种信号途径不可或缺的组成部分,其在植物体内的浓度会随着环境变化而变化,对植物响应非生物胁迫至关重要[6-7]。先前的研究发现了三大类钙受体:钙调素(calmodulin,CaM)、钙依赖性蛋白激酶(calcium-dependent protein kinase,CDPK)和类钙调磷酸酶B亚基蛋白(calcineurin B-like protein,CBL)[8]。其中,CBL能够与CBL互作蛋白激酶(CBL-interacting protein kinase,CIPK)结合形成CBL-CIPK蛋白复合物,并参与信号传导,使植物能够对盐、干旱、低温等胁迫做出响应[7,9-11]。
在拟南芥中,目前共鉴定出10个编码CBL蛋白的基因(AtCBL1~10)[12],其中,AtCBL1、AtCBL2、AtCBL5、AtCBL9和AtCBL10基因已被证实对非生物胁迫具有响应能力[13-17]。玉米中,ZmCBL3、ZmCBL4、ZmCBL5和ZmCBL8通过与ZmCIPK16互作,响应干旱、高盐等非生物胁迫[18]。此外,过表达OsCBL8能够赋予水稻高温抗性,OsCBL5是水稻盐胁迫的负调控因子[19-20]。研究表明低温处理会提高豌豆中PsCBL的转录水平,但干旱处理对PsCBL的表达没有影响[21]。在杨树中,盐胁迫条件下PeCBL1、PeCBL2、PeCBL4、PeCBL5和PeCBL9基因均会上调表达[22]。这些结果进一步表明,CBL基因能够响应多种非生物胁迫,参与调节植物体内离子稳态,协助植物适应环境胁迫。
1 材料和方法
1.1 材料和生长条件
供试材料选用花生品种‘花育23’。花生种子利用质量分数2% NaClO表面灭菌10 min,在无菌去离子水中萌发至4叶期,选取长势相似的幼苗,移植到水培盒中。材料分2组开展研究:对照组为1/2强度Hoagland溶液,处理组为1/2强度Hoagland溶液+200 mmol/L NaCl。分别在培养0、0.5、1、3、6、12、24 h时对根系进行取样,样品在液氮中速冻后保存于-80 ℃。
1.2 AhCBL基因家族成员的鉴定及理化性质分析
从花生基因组数据库(http:∥peanutgr.fafu.edu.cn/News_chinese.php/)下载花生的全基因组序列及注释文件;从TAIR(https:∥www.arabidopsis.org/)下载拟南芥CBL蛋白序列;在pfam网站(http:∥pfam-legacy.xfam.org/)下载CBL隐马尔科夫模型(PF00036)[29];利用TBtools软件进行BLAST和HMMER搜索,筛选关键结构域,确定AhCBL基因家族成员。
使用在线软件ExPASy(https:∥www.expasy.org/)预测AhCBL家族成员的氨基酸长度、分子量、等电点、脂肪指数和亲水指数等理化特性。在Plant-mPLoc网站(http:∥www.csbio.sjtu.edu.cn/bioinf/plant-multi/)对AhCBL家族成员进行亚细胞定位预测。使用SOPMA在线软件(https:∥npsa-prabi.ibcp.fr/)对AhCBL的蛋白二级结构域进行分析与数据统计。
1.3 AhCBL蛋白系统发育树的构建
根据Zhang等[30]的研究,选择拟南芥的10个CBL家族成员和蒺藜苜蓿的13个CBL家族成员,使用TBtools软件对花生、拟南芥和蒺藜苜蓿的CBL蛋白进行多序列比对,使用最大似然法(maximum likelihood,ML)构建系统发育树,bootstrap值为1 000。
1.4 AhCBL基因家族的保守基序和基因结构分析
使用MEME网站(https:∥meme-suite.org/meme/)对AhCBL家族蛋白的保守基序进行预测。使用TBtools软件对AhCBL家族的保守基序及基因内含子、外显子结构进行可视化分析。
1.5 AhCBL基因家族的共线性分析
使用TBtools软件对AhCBL基因进行染色体定位,并使用TBtools中的One Step MCScanX对AhCBL组内以及花生和蒺藜苜蓿CBL组间进行共线性分析,参数设置取默认值。
1.6 AhCBL基因家族的顺式作用元件分析
为分析AhCBL基因家族启动子序列中可能存在的顺式作用元件,提取AhCBL基因上游2 000 bp的DNA序列,使用PlantCARE在线网站(https:∥bioinformatics.psb.ugent.be/webtools/plantcare/html/)进行启动子顺式作用元件预测,筛选出候选的结合元件,并使用TBtools进行可视化分析。
1.7 AhCBL基因在不同组织中的表达模式分析
基于花生基因组数据库下载的AhCBL基因的RNA-seq数据[31],分析AhCBL基因在不同组织的表达模式,包括果皮(Ⅰ、Ⅱ、Ⅲ)、胚(Ⅰ、Ⅱ、Ⅲ、Ⅳ)、外种皮(Ⅰ、Ⅱ)、子叶、花、叶、雌蕊柄、根瘤、根、根和茎、根尖、茎、茎尖等19个组织。基于TBtools软件绘制FPKM表达谱,采用lg 2与row scale标准化。
1.8 荧光定量PCR分析
表1 RT-qPCR特异性引物Tab.1 The specific primers used for RT-qPCR |
| 基因名称 | 正向引物(5'-3') | 反向引物(5'-3') |
|---|---|---|
| TUA5 | CTGATGTCGCTGTGCTCTTGG | CTGTTGAGGTTGGTGTAGGTAGG |
| AhCBL4 | CCGAATTCTCGCTCAACCCT | GCACGAGGACTAAAGACGGA |
| AhCBL33 | TTCACCGGAGGCACTTTCTC | ACTGAGCAATGAAAGGCGGA |
| AhCBL46.1 | TTATGGCCCCGAAGCAGATG | GGCCATGGATCAGAGTCGAA |
| AhCBL61.3 | ATTCACTGACGTGGTTGGCA | GGCGGCACTCCACTAAGTAA |
| AhCBL66 | TGAGGTTGATGCTGATGGCA | ACATGTCTCAACTCACTGGCT |
2 结果与分析
2.1 AhCBL基因家族成员鉴定及理化性质分析
花生基因组中共鉴定出89个CBL基因(表2),基因根据其所在的染色体定位进行命名。这些基因编码的氨基酸数量介于54~774 aa,分子量介于6.429~87.948 kDa,等电点介于4.00~9.34,酸性氨基酸占比95%以上,脂肪系数介于69.40~97.91,亲水指数介于-0.665~0.015,亲水性占比97%以上。亚细胞定位预测结果显示,31个AhCBL蛋白定位于细胞膜,43个AhCBL蛋白定位于细胞核,4个AhCBL蛋白定位于液泡,10个AhCBL蛋白定位于细胞膜和细胞质,1个AhCBL蛋白定位于细胞膜和细胞核。使用SOPMA在线软件对AhCBL蛋白进行二级结构预测,发现AhCBL蛋白的二级结构主要以α螺旋(25.93%~64.43%)和无规则卷曲(7.43%~44.36%)为主,此外,还包括延伸链(2.41%~33.33%)和β折叠(4.82%~18.52%)。
表2 AhCBL基因家族的理化性质Tab.2 Physicochemical properties of the AhCBL gene family |
| 基因名称 | 基因ID | 氨基酸数量/aa | 分子量/kDa | 等电点 | 脂肪系数 | 亲水指数 | 亚细胞定位 |
|---|---|---|---|---|---|---|---|
| AhCBL1 | AH01G10260.1 | 166 | 18.403 | 4.73 | 79.52 | -0.445 | 液泡 |
| AhCBL2 | AH01G11240.1 | 517 | 57.820 | 5.69 | 93.33 | -0.241 | 细胞核 |
| AhCBL3 | AH01G13870.1 | 213 | 24.301 | 4.64 | 94.23 | -0.076 | 细胞膜 |
| AhCBL4 | AH01G23540.1 | 175 | 20.308 | 4.54 | 84.00 | -0.170 | 细胞膜 |
| AhCBL5.1 | AH01G29930.1 | 206 | 23.437 | 4.66 | 92.23 | -0.245 | 细胞膜 |
| AhCBL5.2 | AH01G29930.2 | 226 | 25.834 | 4.73 | 90.13 | -0.291 | 细胞膜 |
| AhCBL6 | AH01G29950.1 | 83 | 9.526 | 4.81 | 89.16 | -0.378 | 细胞核 |
| AhCBL7 | AH01G34550.1 | 553 | 62.594 | 5.96 | 87.40 | -0.425 | 细胞核 |
| AhCBL8.1 | AH02G13470.1 | 584 | 65.618 | 5.24 | 83.65 | -0.380 | 细胞核 |
| AhCBL8.2 | AH02G13470.2 | 584 | 65.618 | 5.24 | 83.65 | -0.380 | 细胞核 |
| AhCBL8.3 | AH02G13470.3 | 584 | 65.618 | 5.24 | 83.65 | -0.380 | 细胞核 |
| AhCBL9 | AH02G22400.1 | 536 | 60.128 | 6.64 | 85.69 | -0.368 | 细胞核 |
| AhCBL10 | AH03G04010.1 | 547 | 62.445 | 6.23 | 77.13 | -0.544 | 细胞核 |
| AhCBL11 | AH03G26220.1 | 284 | 32.668 | 6.47 | 87.57 | -0.366 | 细胞膜 |
| AhCBL12 | AH03G26230.1 | 216 | 24.695 | 5.45 | 93.33 | -0.114 | 细胞膜 |
| AhCBL13 | AH04G00420.1 | 553 | 61.549 | 5.39 | 75.86 | -0.568 | 细胞核 |
| AhCBL14 | AH04G04110.1 | 535 | 59.909 | 5.91 | 74.95 | -0.489 | 细胞核 |
| AhCBL15 | AH04G19890.1 | 540 | 60.550 | 6.04 | 82.54 | -0.421 | 细胞核 |
| AhCBL16 | AH05G03340.1 | 215 | 24.520 | 4.83 | 91.53 | -0.103 | 细胞膜 |
| AhCBL17.1 | AH05G32230.1 | 546 | 61.538 | 6.18 | 83.39 | -0.425 | 细胞核 |
| AhCBL17.2 | AH05G32230.2 | 547 | 61.669 | 6.18 | 83.24 | -0.421 | 细胞核 |
| AhCBL18 | AH05G33410.1 | 157 | 17.282 | 4.38 | 87.13 | -0.207 | 液泡 |
| AhCBL19 | AH06G01890.1 | 538 | 60.818 | 6.50 | 81.52 | -0.511 | 细胞核 |
| AhCBL20 | AH06G11630.1 | 149 | 16.915 | 4.10 | 77.32 | -0.409 | 细胞膜和细胞质 |
| AhCBL21 | AH06G13470.1 | 763 | 86.576 | 8.38 | 80.01 | -0.506 | 细胞核 |
| AhCBL22 | AH06G19880.1 | 149 | 16.861 | 4.12 | 69.40 | -0.619 | 细胞膜和细胞质 |
| AhCBL23 | AH06G27470.1 | 175 | 20.253 | 4.58 | 81.20 | -0.089 | 细胞膜 |
| AhCBL24 | AH07G01500.1 | 226 | 26.119 | 4.81 | 94.91 | -0.245 | 细胞膜 |
| AhCBL25.1 | AH07G03340.1 | 559 | 63.725 | 6.46 | 75.37 | -0.665 | 细胞核 |
| AhCBL25.2 | AH07G03340.3 | 559 | 63.725 | 6.46 | 75.37 | -0.665 | 细胞核 |
| AhCBL26 | AH07G05100.1 | 534 | 59.623 | 5.63 | 77.28 | -0.501 | 细胞核 |
| AhCBL27 | AH07G05500.1 | 149 | 16.861 | 4.12 | 69.40 | -0.619 | 细胞膜和细胞质 |
| AhCBL28 | AH07G05510.1 | 149 | 16.861 | 4.12 | 69.40 | -0.619 | 细胞膜和细胞质 |
| AhCBL29.1 | AH08G04290.1 | 216 | 24.756 | 4.96 | 96.53 | -0.204 | 细胞膜 |
| AhCBL29.2 | AH08G04290.2 | 216 | 24.756 | 4.96 | 96.53 | -0.204 | 细胞膜 |
| AhCBL30 | AH08G28860.1 | 371 | 42.550 | 5.05 | 92.70 | -0.259 | 细胞膜和细胞核 |
| AhCBL31 | AH09G05080.1 | 266 | 30.706 | 4.83 | 96.73 | -0.179 | 细胞膜 |
| AhCBL32 | AH09G19680.1 | 151 | 17.128 | 4.00 | 82.65 | -0.462 | 细胞膜和细胞质 |
| AhCBL33 | AH11G10850.1 | 764 | 85.304 | 8.34 | 96.58 | -0.006 | 细胞核 |
| AhCBL34 | AH11G14310.1 | 213 | 24.373 | 4.60 | 94.23 | -0.090 | 细胞膜 |
| AhCBL35 | AH11G22340.1 | 553 | 62.584 | 5.96 | 86.69 | -0.427 | 细胞核 |
| AhCBL36.1 | AH11G26940.1 | 226 | 25.834 | 4.73 | 90.13 | -0.291 | 细胞膜 |
| AhCBL36.2 | AH11G26940.2 | 226 | 25.834 | 4.73 | 90.13 | -0.291 | 细胞膜 |
| AhCBL37 | AH11G34320.1 | 175 | 20.308 | 4.54 | 84.00 | -0.170 | 细胞膜 |
| AhCBL38 | AH12G16690.1 | 249 | 28.414 | 4.59 | 86.99 | -0.340 | 细胞膜 |
| AhCBL39 | AH12G24630.1 | 536 | 60.157 | 6.20 | 85.52 | -0.371 | 细胞核 |
| AhCBL40 | AH12G33910.1 | 626 | 68.219 | 5.48 | 80.89 | -0.352 | 细胞核 |
| AhCBL41 | AH12G34040.1 | 665 | 73.218 | 5.31 | 78.96 | -0.446 | 细胞核 |
| AhCBL42 | AH13G05160.1 | 538 | 60.157 | 5.69 | 76.17 | -0.545 | 细胞核 |
| AhCBL43 | AH13G05950.1 | 547 | 62.493 | 6.23 | 76.23 | -0.551 | 细胞核 |
| AhCBL44.1 | AH13G32000.1 | 216 | 24.792 | 5.61 | 86.57 | -0.192 | 细胞膜 |
| AhCBL44.2 | AH13G32000.2 | 216 | 24.792 | 5.61 | 86.57 | -0.192 | 细胞膜 |
| AhCBL45 | AH13G32010.1 | 162 | 18.269 | 5.09 | 96.36 | -0.081 | 细胞膜 |
| AhCBL46.1 | AH13G60090.1 | 559 | 62.975 | 5.58 | 82.70 | -0.458 | 细胞核 |
| AhCBL46.2 | AH13G60090.2 | 559 | 62.975 | 5.58 | 82.70 | -0.458 | 细胞核 |
| AhCBL46.3 | AH13G60090.3 | 559 | 62.975 | 5.58 | 82.70 | -0.458 | 细胞核 |
| AhCBL46.4 | AH13G60090.4 | 559 | 62.975 | 5.58 | 82.70 | -0.458 | 细胞核 |
| AhCBL46.5 | AH13G60090.5 | 559 | 62.975 | 5.58 | 82.70 | -0.458 | 细胞核 |
| AhCBL46.6 | AH13G60090.6 | 559 | 62.975 | 5.58 | 82.70 | -0.458 | 细胞核 |
| AhCBL47.1 | AH14G05090.1 | 529 | 59.145 | 5.78 | 76.35 | -0.470 | 细胞核 |
| AhCBL47.2 | AH14G05090.2 | 529 | 59.145 | 5.78 | 76.35 | -0.470 | 细胞核 |
| AhCBL48 | AH14G07260.1 | 231 | 26.595 | 5.31 | 85.67 | -0.310 | 细胞膜 |
| AhCBL49 | AH14G22710.1 | 540 | 60.563 | 6.04 | 83.26 | -0.411 | 细胞核 |
| AhCBL50 | AH15G19030.1 | 157 | 17.308 | 4.46 | 87.13 | -0.218 | 液泡 |
| AhCBL51 | AH15G20870.1 | 547 | 61.635 | 6.08 | 83.58 | -0.401 | 细胞核 |
| AhCBL52 | AH16G00240.1 | 774 | 87.948 | 8.25 | 78.50 | -0.545 | 细胞核 |
| AhCBL53 | AH16G01700.1 | 149 | 16.915 | 4.10 | 77.32 | -0.409 | 细胞膜和细胞质 |
| AhCBL54 | AH16G04360.1 | 538 | 60.790 | 6.50 | 81.52 | -0.510 | 细胞核 |
| AhCBL55 | AH16G24820.1 | 149 | 16.861 | 4.12 | 69.40 | -0.619 | 细胞膜和细胞质 |
| AhCBL56 | AH16G34060.1 | 54 | 6.429 | 4.57 | 86.48 | -0.198 | 细胞膜 |
| AhCBL57 | AH16G38340.3 | 234 | 26.860 | 4.66 | 92.09 | -0.124 | 细胞膜 |
| AhCBL58.1 | AH16G38340.1 | 243 | 27.974 | 4.69 | 93.13 | -0.132 | 细胞膜 |
| AhCBL58.2 | AH16G38340.2 | 227 | 26.035 | 4.53 | 97.49 | 0.011 | 细胞膜 |
| AhCBL59.1 | AH16G44040.1 | 248 | 28.904 | 4.68 | 90.85 | -0.156 | 细胞膜 |
| AhCBL59.2 | AH16G44040.2 | 248 | 28.904 | 4.68 | 90.85 | -0.156 | 细胞膜 |
| AhCBL60 | AH16G46940.1 | 249 | 28.314 | 9.34 | 97.91 | 0.015 | 细胞膜 |
| AhCBL61.1 | AH17G00410.1 | 560 | 63.090 | 5.58 | 81.86 | -0.477 | 细胞核 |
| AhCBL61.2 | AH17G00410.2 | 560 | 63.090 | 5.58 | 81.86 | -0.477 | 细胞核 |
| AhCBL61.3 | AH17G00410.3 | 560 | 63.090 | 5.58 | 81.86 | -0.477 | 细胞核 |
| AhCBL62 | AH17G01640.1 | 226 | 26.092 | 4.81 | 94.91 | -0.233 | 细胞膜 |
| AhCBL63 | AH17G04170.1 | 149 | 16.861 | 4.12 | 69.40 | -0.619 | 细胞膜和细胞质 |
| AhCBL64 | AH17G04180.1 | 149 | 16.861 | 4.12 | 69.40 | -0.619 | 细胞膜和细胞质 |
| AhCBL65.1 | AH17G27800.1 | 223 | 25.444 | 4.98 | 96.55 | -0.173 | 细胞膜 |
| AhCBL65.2 | AH17G27800.2 | 216 | 24.754 | 5.04 | 97.87 | -0.168 | 细胞膜 |
| AhCBL66 | AH19G24870.1 | 151 | 17.128 | 4.00 | 82.65 | -0.462 | 细胞膜和细胞质 |
| AhCBL67 | AH20G01640.1 | 166 | 18.376 | 4.73 | 79.52 | -0.429 | 液泡 |
| AhCBL68 | AH20G34090.1 | 580 | 64.953 | 5.20 | 79.50 | -0.354 | 细胞核 |
| AhCBL69.1 | AH00G03010.1 | 554 | 63.115 | 6.46 | 76.05 | -0.635 | 细胞核 |
| AhCBL69.2 | AH00G03010.2 | 554 | 63.115 | 6.46 | 76.05 | -0.635 | 细胞核 |
2.2 花生、蒺藜苜蓿和拟南芥CBL家族系统发育树分析
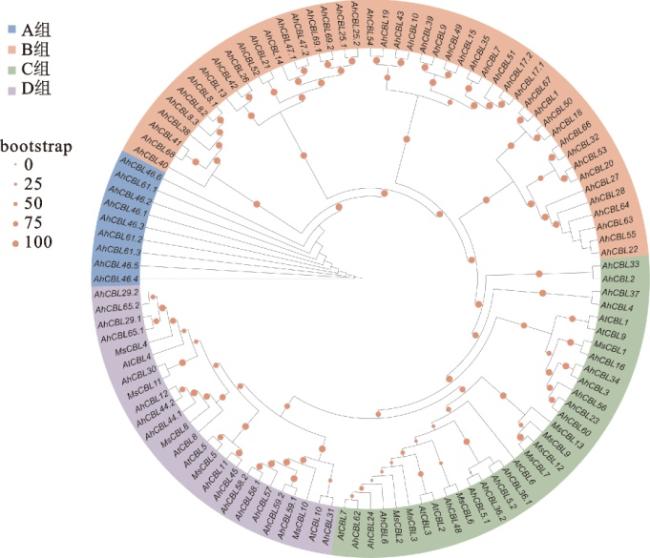
对花生、蒺藜苜蓿和拟南芥中共112个CBL蛋白构建系统发育树,将花生中的CBL蛋白根据主要分支分为A、B、C、D 4组(图1)。其中,B组最多,有46个成员;A组最少,有9个成员;C组和D组分别有18个和16个成员。
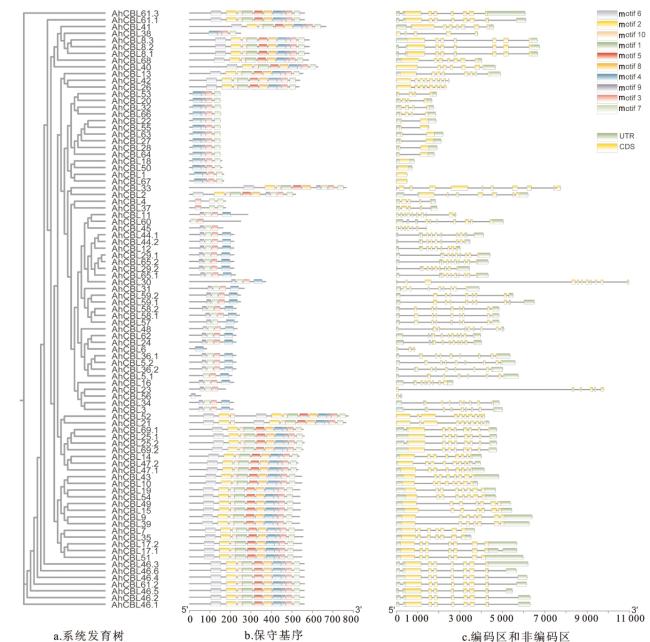
2.3 AhCBL家族保守基序和基因结构分析
2.4 AhCBL基因组内和组间共线性分析
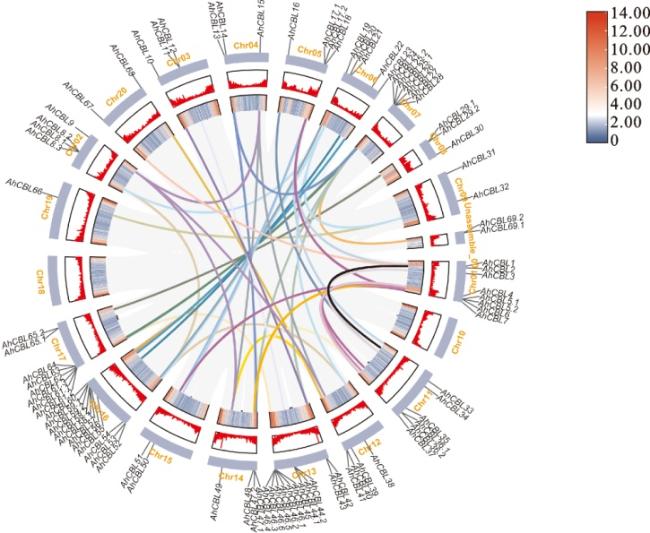
89个AhCBL基因共分布在19条染色体上,其中,13号染色体上最多,有11个AhCBL基因,19号染色体上只有AhCBL66。AhCBL组内共线性分析结果表明, 89个AhCBL成员之间存在47组共线性关系(图3)。
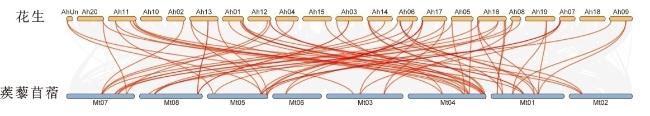
由图4可以看出,花生与蒺藜苜蓿CBL基因之间存在70组共线性。其中,16号染色体上的共线性对最多,有7组;6、11、17号染色体次之,有6组;2号和15号染色体上的共线性对最少,分别都只有1组。蒺藜苜蓿4号染色体上分别有4个CBL与花生1号染色体和11号染色体上的AhCBL发生配对,蒺藜苜蓿1号染色体上有4个CBL与花生16号染色体上的AhCBL发生配对,说明蒺藜苜蓿4号染色体与花生1号和11号染色体、蒺藜苜蓿1号染色体与花生16号染色体在CBL进化的线性关系上高度一致。
2.5 AhCBL家族顺式作用元件分析
通过在线软件PlantCARE对AhCBL基因家族上游2 000 bp的启动子序列进行顺式作用元件分析,进而预测各基因潜在的生物学功能。在启动子区域中共发现18个顺式作用元件,主要包括生长发育、激素响应和胁迫响应三大类元件(图5)。其中,胁迫响应元件包括厌氧响应、低温响应、防御与应激响应元件,激素响应元件主要包括赤霉素、水杨酸和茉莉酸甲酯响应元件等。以上结果表明AhCBL可能参与激素响应和非生物胁迫应答等过程。

不同AhCBL预测到不同的顺式作用元件,例如AhCBL4、AhCBL5.1、AhCBL5.2、AhCBL9、AhCBL13、AhCBL14、AhCBL16、AhCBL19、AhCBL23、AhCBL24、AhCBL27、AhCBL33、AhCBL36.1、AhCBL36.2、AhCBL38、AhCBL39、AhCBL41、AhCBL47.1、AhCBL47.2、AhCBL51、AhCBL54、AhCBL55、AhCBL56、AhCBL57、AhCBL58.1、AhCBL58.2、AhCBL62、AhCBL63、AhCBL68的启动子区域中含有低温响应相关顺式作用元件;AhCBL1、AhCBL2、AhCBL3、AhCBL8.2、AhCBL8.3、AhCBL9、AhCBL10、AhCBL14、AhCBL16、AhCBL20、AhCBL22、AhCBL25.1、AhCBL25.2、AhCBL28、AhCBL29.1、AhCBL29.2、AhCBL32、AhCBL34、AhCBL35、AhCBL36.1、AhCBL38、AhCBL39、AhCBL40、AhCBL41、AhCBL43、AhCBL47.1、AhCBL48、AhCBL50、AhCBL54、AhCBL55、AhCBL65.1、AhCBL65.2的启动子区域含有防御与应激响应元件;除AhCBL4、AhCBL10、AhCBL11、AhCBL13、AhCBL19、AhCBL20、AhCBL22、AhCBL29.1、AhCBL29.2、AhCBL32、AhCBL41、AhCBL45、AhCBL46.1、AhCBL46.2、AhCBL46.3、AhCBL46.4、AhCBL46.5、AhCBL46.6、AhCBL53、AhCBL54、AhCBL55、AhCBL65.1、AhCBL65.2、AhCBL68、AhCBL69.1、AhCBL69.2外的其余63个AhCBL启动子区域都含有厌氧响应元件。可见,AhCBL在响应逆境胁迫中可能发挥重要作用。
2.6 AhCBL基因组织表达模式分析
根据花生基因组数据库网站下载得到的各组织表达量进行AhCBL基因表达模式分析,相对转录本水平取lg 2(FPKM+1)。结果显示(图6),31个AhCBL基因(34.8%)在花生叶片中高表达,34个AhCBL基因(38.2%)在花生茎部高表达,43个AhCBL基因(48.3%)在花生根部高表达,而叶片、茎部和根部分别有3个、5个和18个家族成员不表达,其中AhCBL10和AhCBL18仅在叶片中表达。同时可以看到,AhCBL22、AhCBL27、AhCBL28、AhCBL55、AhCBL63、AhCBL64这6个基因在各组织中均高表达。
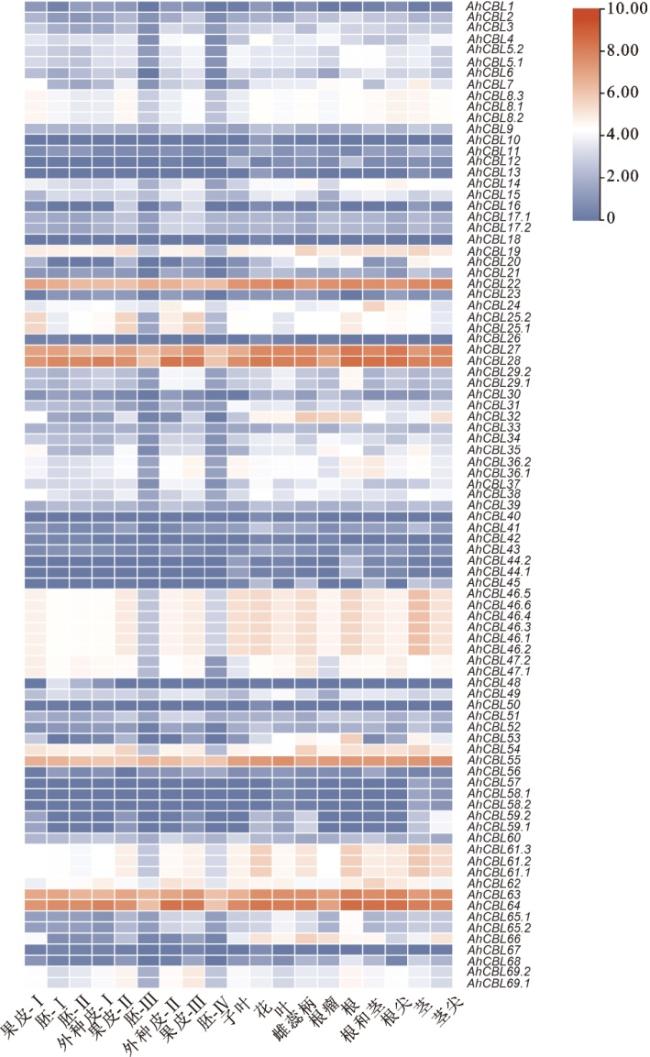
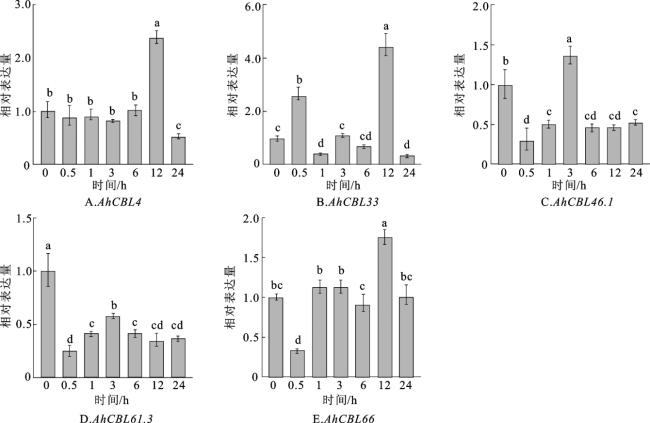
2.7 盐胁迫下AhCBL基因表达模式分析
为研究AhCBL基因对盐胁迫的响应,选取在根部表达量较高的5个AhCBL基因(AhCBL4、AhCBL33、AhCBL46.1、AhCBL61.3、AhCBL66)进行RT-qPCR验证。结果显示(图7),在200 mmol/L NaCl盐胁迫处理12 h后,AhCBL4、AhCBL33和AhCBL66的表达量均显著升高,AhCBL46.1在处理3 h时表达量显著升高,而AhCBL61.3对盐胁迫表现出负响应。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 讨论
植物中的Ca2+感知蛋白感知并响应Ca2+浓度的变化,调控特定的信号转导并参与植物对各种环境刺激(包括生物和非生物胁迫)的响应,在植物的各种生命活动中发挥重要作用[33]。作为Ca2+感知蛋白,CBL蛋白通过结合Ca2+并与特定蛋白激酶相互作用来传递钙信号。目前,CBL家族成员已在多种植物中被鉴定,如苜蓿(68个)[30]、小麦(68个)[34]、水稻(10个)[35]和玉米(12个)[36]。本研究利用生物信息学分析,在花生中共鉴定出89个CBL基因家族成员,89个AhCBL基因编码的氨基酸数在54~774 aa之间,相对分子质量介于6.429~87.948 kDa,等电点介于4.00~9.34,AhCBL蛋白中的酸性氨基酸占比95%以上,脂肪系数介于69.40~97.91。已有许多研究显示,外显子和内含子的结构在基因家族的演化和成员功能分化中起着关键作用。内含子的数量和长度对真核生物的基因表达也具有至关重要的调控作用[37-39]。本研究发现,AhCBL家族内各成员的内含子数量介于0~10个,且内含子长度差异较大,暗示AhCBL基因家族组内成员之间的基因功能差异较大(图 2c)。花生CBL基因在染色体上的分布呈不规则状态,在13号染色体上的分布较为密集(图3),由此推测花生基因组中的AhCBL之间可能存在功能冗余。在逆境条件下,机体可通过激活转录因子并结合基因的顺式作用元件来调节基因表达,以适应外界压力。研究指出,CBL基因家族普遍参与植物的发育过程以及对非生物胁迫的响应[40-41]。本研究通过分析顺式作用元件发现,AhCBL家族成员基因上游2 000 bp启动子区域含有大量与抗逆、激素响应和生长发育相关的结合元件(图 5)。此外,AhCBL基因在花生的不同组织中广泛表达,包括根、成熟叶、发育中的花以及固氮根瘤(图 6),表明它们在植物的生长发育过程中扮演着关键角色。
高盐度是限制植物生长发育和作物产量的主要环境因素,CBL和CIPK基因家族在植物应对盐胁迫中具有重要作用[42-43]。通常情况下,根是植物感知和应对胁迫的关键器官。在拟南芥中,AtCBL4/SOS3激活AtCIPK24/SOS2,启动SOS途径,以维持盐胁迫下根系中的离子平衡[44-45]。这一途径在水稻、玉米和甘蔗的盐胁迫反应中也被广泛报道[18,20,46-48]。在大豆中共鉴定出15个CBL基因家族成员,其中GmCBL1基因在大豆对盐和碱胁迫的响应中发挥重要作用。GmCBL1可能作为钙信号感受器,感知细胞内钙离子浓度的变化,并激活下游CBL-CIPK信号通路,该通路进而调控离子通道、渗透调节、抗氧化等基因的表达,增强植物对盐、碱胁迫的耐受性[49]。苜蓿中多数CBL基因在盐胁迫和干旱胁迫条件下表达水平显著上调,表明它们可能在苜蓿抗逆性调控中发挥重要作用[30]。鹰嘴豆中CaCBL1、CaCBL4、CaCBL10等基因在根、茎、叶等组织中均有表达,且在盐胁迫、干旱胁迫下表达量显著上调,表明它们参与鹰嘴豆的非生物胁迫响应[50]。利用盐、干旱、寒冷、损伤和脱落酸暴露等多种刺激均可诱导AtCBL1表达[51-52]。同时,AtCBL9在负向调节冷胁迫的过程中,使得拟南芥对脱落酸不敏感[53]。在转基因拟南芥中,紫花针茅CBL6的过表达提高了植株的耐寒性并降低了耐旱性,萝卜的BrrCBL9.2则提高了植株的耐盐性[54-55]。AhCBL基因在不同亚家族中的表达模式各不相同。本研究中,在盐胁迫条件下,AhCBL4、AhCBL33、AhCBL46.1和AhCBL66的表达均上调,而AhCBL61.3的表达受到抑制(图 7)。这些结果表明,AhCBL4、AhCBL33、AhCBL46.1和AhCBL66能够提高花生的耐盐性,而AhCBL61.3可能具有相反的功能。
4 结论
本研究在花生中共鉴定出89个CBL基因。根据系统进化和结构域分析结果,花生CBL基因家族可分为4组。基因表达分析显示,AhCBL4、AhCBL33、AhCBL46.1、AhCBL61.3和AhCBL66在花生的不同组织中均表现出显著的调节作用。RT-qPCR结果表明,AhCBL4、AhCBL33、AhCBL46.1和AhCBL66可能对盐胁迫具有正向调节作用,而AhCBL61.3可能对盐胁迫具有负向调控作用。研究结果为今后研究花生中CBL基因的生物学功能奠定了基础。