

菱形(Rhomboid)蛋白属于丝氨酸蛋白酶家族,是广泛存在于各种生物中的保守性膜内蛋白;菱形蛋白表现出多种底物特异性,可通过识别结合不同的底物,在植物生长发育调节方面发挥重要作用[1-2]。杨洋[3]在腊梅(Chimonanthus praecox L.)中初步探究了CpRhomboid基因的功能表达,发现 CpRhomboid基因对植物的耐盐性不产生影响,可能通过调节MAPKK基因的表达进而参与植物的抗旱性调节;赵莹[4]在大白菜(Brassica rapa L.)中鉴定出1个可能影响雄性不育的基因BrRBL3,其与Rhomboid同源,在花粉中表达量最高,该基因异源表达可使雄性不育的拟南芥(Arabidopsis thaliana)花粉正常发育;Knopf等[5]发现拟南芥AtRBL6可能响应非生物胁迫。此外,有研究表明,在幼苗期冷处理可以诱导AtRBL10,说明AtRBL10可以响应冷胁迫[6];也有研究显示,AtRBL10的表达量在热胁迫后上调,其表达相关基因中有29%响应热胁迫[5]。目前,关于菱形蛋白的研究主要集中在医学领域[7⇓⇓-10],在植物研究中少有报道,其响应非生物胁迫的机制尚不清楚,亟需进一步研究和探索。
1 材料与方法
1.1 材料
将保存于4 ℃冰箱的青杄种子用赤霉素(gibberellins,GA)溶液浸泡24 h,使用蒸馏水多次润洗。将润洗干净的种子分散在放有湿润滤纸的玻璃培养皿中,置于21 ℃的光照培养箱中培养,光照16 h、黑暗8 h,始终保持水分充足。待种子萌发后移栽至育苗容器中(m营养土∶m蛭石=1∶1),放置于温室生长,定期浇水防虫。
所用野生型拟南芥为哥伦比亚野生型(wild-type,WT)。拟南芥种子先用75%的酒精处理5 min,再用5%的次氯酸钠溶液处理15 min,接着使用灭菌蒸馏水反复润洗。将种子播在MS固体培养基上,置于4 ℃黑暗环境培养3 d,取出后放在光照培养箱中继续培养,待长出4片真叶后,将幼苗移栽至育苗盆中生长,定期浇水防虫。
所用烟草为本氏烟草。将烟草种子撒播在混合土(m营养土∶m蛭石∶m珍珠岩=2∶1∶1)中,覆膜放置于温室,3 d后揭膜正常生长,约2周后将幼苗单株移栽,定期浇水防虫。
所用马铃薯组培苗为银川市农林科学院赠送,品种为“大西洋”。
1.2 方法
1.2.1 载体构建及转基因植株培养
使用限制性内切酶Sma Ⅰ和EcoR Ⅰ酶切pCM1205载体,纯化回收后,使用T4连接酶将酶切产物与目的基因PwRhomboid过夜连接,将连接好的PwRhomboid-pCM1205重组载体转化根癌农杆菌感受态GV3101中,保存备用。
配置pCM1205农杆菌浸染液,在超净工作台中将马铃薯切块,放入浸染液中,摇晃8 min,取出马铃薯小块,放置于滤纸上吸除表面多余菌液,置于MS固体培养基(含100 μmol/L 乙酰丁香酮)中黑暗培养2 d。然后,使用300 mg/L 头孢霉素水和无菌水洗菌后,放入筛选培养基(1/2 MS+1 mg/L萘乙酸+250 mg/L头孢霉素)中生长。取部分叶片及茎段提取RNA,反转录为cDNA,通过RT-qPCR鉴定阳性植株。马铃薯阳性植株的鉴定参照李兴芬[18]的方法。
研究所用引物序列信息见表1。
表1 引物序列信息Tab.1 Information of primer sequences |
| 用途 | 引物名称 | 引物序列(5'-3') |
|---|---|---|
| RT-qPCR | AtActin-F | GGTAACATTGTGCTCAGTGGTGG |
| AtActin-R | AACGACCTTAATCTTCATGCTGC | |
| PwEF1α-RT-F | AACTGGAGAAGGAACCCAAG | |
| PwEF1α-RT-R | AACGACCCAATGGAGGATAC | |
| PwRhomboid-RT-F | GGAGAAATGGGGAAGCATGTG | |
| PwRhomboid-RT-R | GCAAAGAAGCCTCTACCCATTAAC | |
| 载体构建 | pEASY-T1-PwRhomboid-F | GGCGATTCCTTGCGTGTG |
| pEASY-T1-PwRhomboid-R | CTGCAAAGAAGCCTCTACCCATTA | |
| pCM1205-PwRhomboid-F | CGCGGATCCATGGGGAAGCATGTGCAAATG | |
| pCM1205-PwRhomboid-R | TCCCCCGGGTTAACTGCAGAGTTGGGAACAAAG |
1.2.2 PwRhomboid的表达特性分析
选用温室中生长状况良好、约8周的青杄幼苗,进行非生物胁迫处理和激素处理,包括干旱处理、低温处理(4 ℃)、高温处理(42 ℃)、盐胁迫处理(200 mmol/L NaCl)、脱落酸(abscisic acid,ABA)处理(100 μmol/L ABA)、赤霉素处理(400 μmol/L GA)、茉莉酸甲酯(methyl jasmonate,MeJA)处理(100 μmol/L MeJA)和水杨酸(salicylic acid,SA)处理(500 μmol/L SA)。分别在处理0、3、6、12 h时取样,以无菌水处理的幼苗作为空白对照,分析各组中PwRhomboid基因在根、茎、幼嫩叶、成熟叶、花粉和种子中的表达情况,每组生物学重复3次。提取青杄幼苗RNA,反转录为cDNA,通过RT-qPCR测定PwRhomboid的表达情况,选择PwEF1α为内参基因。
1.2.3 亚细胞定位
将含有PwRhomboid-pCM1205重组载体的农杆菌活化,用枪头蘸取部分菌加入液体抗性培养基中,在28 ℃、200 r/min的摇床中培养至OD值为0.8。菌液在5 000 r/min离心机中离心10 min集菌,使用烟草缓冲液重悬菌体,调OD值为0.5,静置2 h后注射烟草叶片,正常生长2 d后用激光共聚焦显微镜在激发光488 nm条件下观察转化的烟草叶片,并拍照保留。
1.2.4 拟南芥和马铃薯干旱表型实验
拟南芥幼苗表型:将WT、pCM1205、PwRhomboid-L1、PwRhomboid-L2种子分别播在含有不同物质的量浓度(0、100、200、300 mmol/L)甘露醇的MS固体培养基上,4 ℃黑暗环境春化3 d。取出春化后的种子转移到光照培养箱中培养,每天观察种子的萌发情况并记录数据[20]。
拟南芥成苗干旱表型:将WT、pCM1205、PwRhomboid-L1、PwRhomboid-L2种子播种在MS固体培养基上,待长出4片幼嫩叶片后,移入土壤中继续生长。进行自然干旱处理,待成苗出现萎蔫干枯状态时,复水3 d,统计存活率并拍照记录。
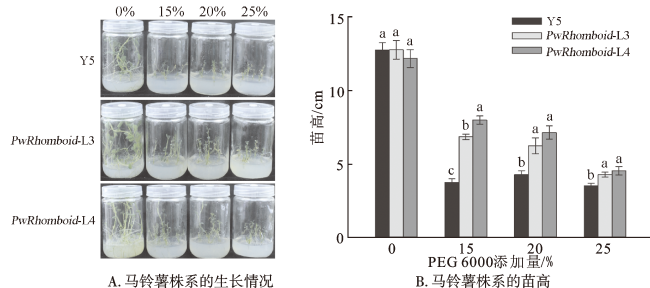
马铃薯组培苗干旱表型:将野生型马铃薯Y5、PwRhomboid-L3、PwRhomboid-L4分别移栽在含有0%、15%、20%和25% PEG6000的MS固体培养基上,进行干旱表型实验,统计植株苗高并拍照记录。
1.2.5 生理指标测定
1.2.6 数据处理
使用SPSS软件对数据进行显著性分析,使用SigmaPlot 14.0软件作图。
2 结果分析
2.1 PwRhomboid多序列比对与进化树分析

2.2 PwRhomboid的表达特性分析
通过PwRhomboid的表达量变化反映温室培育8周且生长状况良好的青杄幼苗对不同非生物胁迫处理(干旱、低温、高温、盐)和激素处理(ABA、GA、SA、MeJA)的响应情况。结果显示,在干旱处理条件下,随着处理时间的增加,PwRhomboid的表达量逐渐升高,处理12 h时其表达量约为对照组的28.7倍(图2A);在低温处理条件下,PwRhomboid的表达量在处理3 h时上调最明显,为对照组的3.5倍(图2B);在高温处理条件下,PwRhomboid的表达量在12 h时达到对照组的1.7倍(图2C);PwRhomboid对盐胁迫无明显响应(图2D)。上述结果说明,PwRhomboid的表达受干旱胁迫诱导最为强烈。
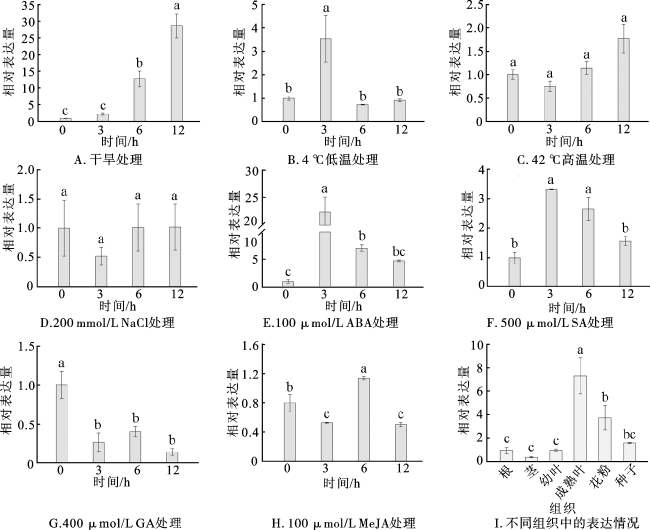
为进一步探究PwRhomboid的功能,使用RT-qPCR测定PwRhomboid在青杄不同部位的表达水平。结果显示,PwRhomboid在成熟叶中的表达量最高,其次是花粉和种子,在茎中的表达量最低(图2I)。
2.3 亚细胞定位
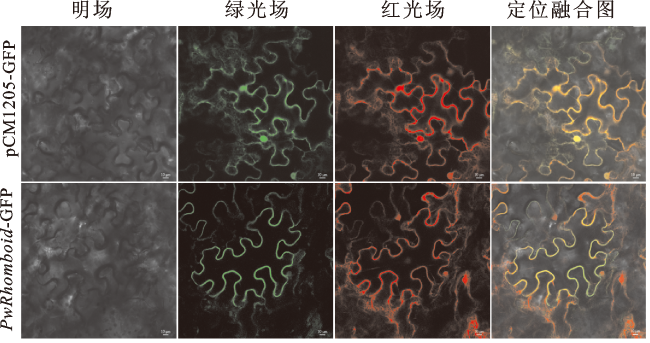
通过瞬时转化烟草叶片对PwRhomboid蛋白进行亚细胞定位分析。将含有pCM1205-GFP和PwRhomboid-GFP表达载体的农杆菌转化进烟草叶片,以pCM1205-GFP为对照组,PwRhomboid-GFP为实验组,观察绿色荧光蛋白(green fluorescent protein,GFP)定位情况,结果如图3所示。可以看出,在对照组的细胞膜、细胞质和细胞核中都有GFP绿色荧光,而在瞬转PwRhomboid-GFP烟草叶片细胞的细胞膜上能观察到GFP荧光,说明PwRhomboid蛋白主要定位于细胞膜上。
2.4 过表达PwRhomboid能够提高干旱胁迫下拟南芥种子的萌发
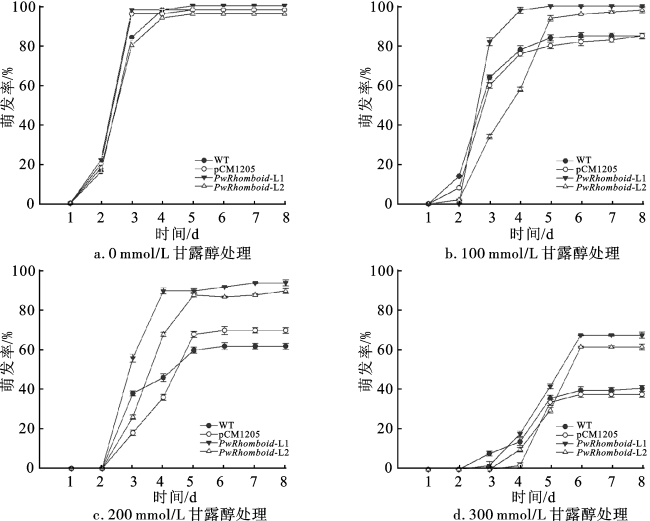
为探究干旱胁迫下PwRhomboid对拟南芥种子萌发的影响,将野生型(WT)、空载株系(pCM1205)和过表达株系(PwRhomboid-L1、PwRhomboid-L2)的拟南芥种子播种在含有不同物质的量浓度(0、100、200、300 mmol/L)甘露醇的MS固体培养基中,连续8 d统计各株系种子的萌发情况。结果表明,在不含甘露醇的培养基中,WT、pCM1205和过表达PwRhomboid的拟南芥种子萌发没有明显差异(图4a);使用100 mmol/L甘露醇处理后,WT、pCM1205种子的萌发率分别降低15.1%、16.4%,而PwRhomboid-L1种子的萌发率没有变化,PwRhomboid-L2种子的萌发率降低1.8%(图4b);使用200 mmol/L甘露醇处理后,WT、pCM1205种子的萌发率分别降低38.2%、30.6%,PwRhomboid-L1、PwRhomboid-L2种子的萌发率分别降低6.3%、9.2%(图4c);使用300 mmol/L甘露醇处理后,所有株系种子的萌发都被抑制,但过表达株系PwRhomboid-L1和PwRhomboid-L2种子的萌发率分别比野生型种子高27.1%和21.3%(图4d)。以上结果表明,过表达PwRhomboid能够提高拟南芥对由甘露醇引起的干旱胁迫的耐受能力。
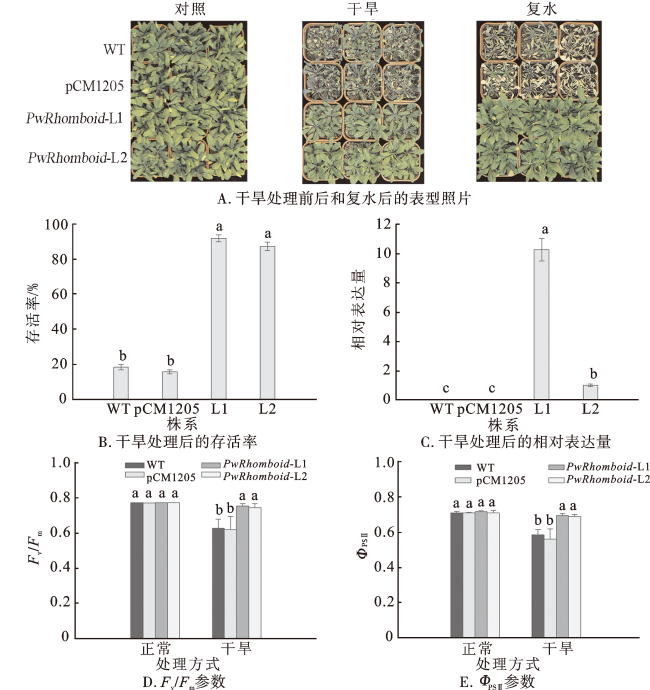
2.5 过表达PwRhomboid能够提高拟南芥成苗的耐旱性
2.6 过表达PwRhomboid能够提高马铃薯的干旱耐受性
为验证PwRhomboid在不同物种中的耐旱功能,将PwRhomboid转化马铃薯,进行干旱处理,将野生型(Y5)和PwRhomboid过表达株系(PwRhomboid-L3、PwRhomboid-L4)的马铃薯继代苗分别接种在含有0%、15%、20%、25% PEG6000的MS培养基中。结果表明,在不添加PEG6000的对照组中,不同株系的生长状态没有明显区别。随着PEG6000添加量的增加,各株系的生长都受到不同程度地抑制。对不同株系的苗高进行统计,发现在15% PEG6000处理下,野生型、PwRhomboid-L3、PwRhomboid-L4的苗高分别为3.74 cm、6.85 cm、7.98 cm;在25% PEG6000处理下,野生型、PwRhomboid-L3、PwRhomboid-L4的苗高分别为3.50 cm、4.27 cm、4.54 cm,PwRhomboid过表达株系的苗高始终比野生型高(图6)。以上结果表明,PwRhomboid能够提高马铃薯对干旱胁迫的抵抗能力,且能在多种物种中发挥植物耐旱性。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 讨论
膜蛋白是细胞膜结构的重要组成部分,其在很大程度上决定着植物的信息传导、物质交换、免疫应答等生命活动[23]。大量研究发现,膜蛋白与植物响应干旱胁迫具有紧密联系[24⇓⇓-27],而菱形蛋白被认为是自然界中分布最广泛的膜蛋白[28]。菱形蛋白是常见的膜内丝氨酸蛋白酶,菱形基因存在于所有生物体中[29]。有研究表明,拟南芥中菱形蛋白AtRBL10能够响应冷胁迫[6]。本研究通过RT-qPCR实验发现PwRhomboid可以响应干旱胁迫,在干旱处理12 h时PwRhomboid基因的表达量升高28.7倍;同时发现PwRhomboid也会响应低温胁迫,说明PwRhomboid可能参与干旱、低温应答途径。植物激素在植物响应干旱胁迫中发挥着重要作用,如ABA、GA等[30⇓-32];在拟南芥中,AtRBL10可能影响茉莉酸信号传导[33]。因此,本研究检测了ABA、GA、SA、MeJA处理对PwRhomboid相对表达量的影响,结果表明PwRhomboid基因对ABA、SA诱导有响应。此外,本研究中PwRhomboid在青杄成熟叶中的表达量最高,在根和茎中表达量较低,推测PwRhomboid基因可能参与调控植物的叶片发育;在腊梅(Chimonanthus praecox L.)中也发现CpRhomboid可以调控植物叶和花的发育[3]。此前,在对拟南芥的相关研究中发现,Rhomboid蛋白的定位不尽相同,AtRBL1和AtRBL2定位于高尔基体,并在所有植物组织中表达[34];而AtRBL8和AtRBL9则定位在叶绿体[35]。本研究发现PwRhomboid蛋白主要定位于细胞膜,其可能参与细胞质膜间的反应。
为验证PwRhomboid在响应植物干旱胁迫中的功能,本研究在拟南芥和马铃薯中转化PwRhomboid并获得过表达植株。经干旱处理后,PwRhomboid过表达拟南芥PwRhomboid-L1和PwRhomboid-L2的生长状况良好,而WT和pCM1205 株系的叶片严重萎蔫;复水后,过表达植株的存活率显著高于WT和pCM1205植株,说明PwRhomboid能够提高植物在干旱胁迫下的存活率。干旱胁迫会影响植物的ROS通路,进而影响植物的光合作用和光化学反应速率[36-37]。最大光化学效率(Fv/Fm)和实际光化学效率(ΦPSⅡ)是使用最广泛的、能够表示光化学反应状况的叶绿素荧光参数。在干旱胁迫下,Fv/Fm、ΦPSⅡ会降低,Fv/Fm、ΦPSⅡ数值越高说明植物光合作用受到胁迫的影响越小,生长状态越好;数值越低说明光合作用受到胁迫的影响越大,生长状态越差[38⇓-40]。本研究的组织定位表达分析显示,PwRhomboid在青杄成熟叶中高表达,推测其通过叶片参与调节响应并提高植物耐旱性。在正常条件下,WT、pCM1205株系和PwRhomboid过表达株系没有明显差异;干旱处理后,WT、pCM1205株系及PwRhomboid过表达株系的Fv/Fm、ΦPSⅡ虽然都有所降低,但WT和pCM1205株系降低更明显,说明过表达PwRhomboid植株受到干旱胁迫的影响更小,对干旱胁迫的抗性更强。使用PEG模拟对转基因马铃薯进行干旱表型实验,发现PwRhomboid过表达株系PwRhomboid-L3、PwRhomboid-L4具有比野生型更高的苗高,表明PwRhomboid能够提高马铃薯对干旱胁迫的耐受能力。在本研究的基础上,后期仍需进一步完善PwRhomboid基因的耐旱表型实验及相关生理指标检测等,深入探索PwRhomboid基因的耐旱机制。
4 结论
本研究发现PwRhomboid基因受到干旱、低温等逆境胁迫及ABA、SA等激素的诱导,PwRhomboid蛋白主要定位于细胞膜,在拟南芥和马铃薯中过表达PwRhomboid能够提高转基因植株的耐旱性。