

随着全球化进程的加快,跨国贸易、旅游业和交通业快速发展,外来生物的引入风险快速增加。我国是深受外来入侵物种危害的国家之一[1]。外来入侵物种不仅会破坏本地的自然生态系统,危害农业生产,还会严重威胁到社会经济的发展,甚至是人类健康[2]。据统计,外来入侵物种在我国每年造成的经济损失超过2 000亿元,其中农业损失占到61.5%[1]。防范外来物种入侵已成为保障国家生态安全的重要一环[3]。我国于2021年4月15日颁布了《中华人民共和国生物安全法》,其中有4个法条直接涉及外来入侵物种的管理,这为防范外来物种入侵提供了法律保障[4]。2022年8月施行的《外来入侵物种管理办法》作出明确规定:任何单位和个人未经批准,不得擅自引进、释放或者丢弃外来物种[5]。习近平同志在党的二十大报告中明确指出“加强生物安全管理,防治外来物种侵害”。由此可见,国家对防范外来物种入侵的重视程度很高。
由于外来入侵物种治理难度大,建立预警机制是阻止外来生物入侵和发生危害最经济有效的措施[6]。准确预测外来物种的潜在入侵区域,可以为其防控和治理提供重要参考依据。生态位模型已被广泛用于预测物种的潜在适生区,其基本原理是以物种已知的分布数据和相关环境因子为基础,按照一定的算法推算出物种的生态位需求,并将计算结果投射到特定的时间和空间,从而获得该物种的适生分布区[7]。应用生态位模型预测入侵物种潜在分布的前提条件是生态位保守性[6],即随着时间和空间的变化,物种保持其祖先生态位不变的倾向[8]。有研究表明,尽管大多数物种的气候生态位具有保守性[9],但仍然有部分入侵物种在到达新生环境后摆脱了其在原产地种群分布的限制因素(竞争、捕食、扩散等),有可能产生适应性的生态位漂移[10]。因此,在模拟物种潜在分布时,要考虑生态位的分化。最大熵模型(MaxEnt)、区域环境模型(DOMAIN)、遗传算法(GARP)、生态位因子分析(ENFA)、生物气候分析系统(BIOCLIM)等是常见的生态位模型。其中,最大熵模型因其预测性能稳定、评价精度高而在外来物种入侵预测[11-12]、濒危物种保护[13]、疾病监测预警[14]等方面得到广泛应用。
多花黑麦草(Lolium multiflorum Lam.)又称意大利黑麦草,是禾本科(Poaceae)黑麦草属(Lolium)的一年生、越年生或短期多年生植物。它原产于欧洲,经引种现已广泛分布于欧洲、美洲和西南亚洲[15]。因多花黑麦草茎叶柔嫩多汁、适口性好且营养丰富,被作为牧草引入我国进行栽培和繁育[16],之后在我国多地广泛种植,其作为一种优质牧草在畜牧养殖和草坪绿化等方面曾有很大贡献。此外,多花黑麦草残茬还能作为绿肥促进农作物水稻的生长[17]。但近些年,由于农田耕作制度的调整、大型机械的推广以及农田管理的疏忽,多花黑麦草在陕西、河南、江苏、四川等地出现混入麦田并造成危害的情况[16]。它不仅会与小麦争夺水、肥、空间,影响小麦长势,还极易出现倒伏,并引起小麦倒伏,对小麦产量造成严重影响。多花黑麦草种子寿命长,叶片光滑狭窄不易着药,且抗药性强,难以从农田里彻底根除[18]。
目前,多花黑麦草的扩散趋势和入侵风险尚不清楚,本文基于多花黑麦草在世界范围的地理分布数据及相关环境因子,先通过环境主成分分析模型对比其原产地和入侵地种群的生态位差异,分析其在入侵中国过程中的生态位进化机制;再基于多花黑麦草在中国、除中国外的全球分布数据分别构建生态位模型,分析其在当前气候下的潜在适生区;最后根据模型结果,分析入侵阶段。通过对多花黑麦草进行入侵风险预警,为其防治提供研究基础。
1 数据与方法
1.1 数据来源
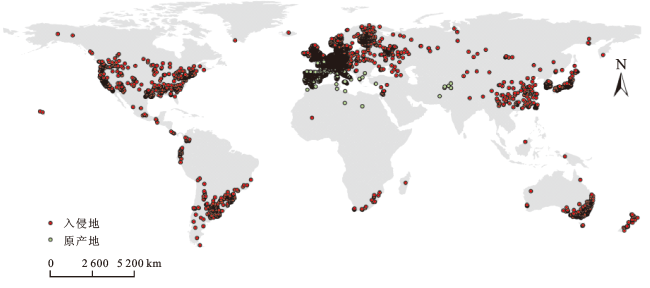
多花黑麦草分布数据主要来源于2个途径:1)从全球生物多样性信息网络(GBIF, http://www.gbif.org/)中共获取其在全球的地理分布数据41 322条[19]。2)通过中国数字植物标本馆(CVH, http://www.cvh.ac.cn/)查询多花黑麦草在中国的分布数据,部分仅有地点描述而没有地理坐标的分布数据在百度地图拾取坐标系统中获取其经纬度坐标,共得到104条分布数据。为减少采样偏差对生态位模型预测的影响,对所得的全部分布点数据删除重复项和冗余点(每2.5'保留1个分布点),并剔除海上分布点,最终得到有效数据13 232条(图1)。其中,原产地分布数据共1 796条,在入侵地中国的分布数据139条。
由于影响物种分布的环境变量主要是气候因素[20],本研究选取了WorldClim数据库(http://www.worldclim.org/)中与温度和降水相关的19个生物气候因子作为环境变量(分辨率为2.5')纳入模型分析。
1.2 研究方法
1.2.1 环境因子筛选
借助ENMTools程序,计算19个环境变量之间的Pearson相关系数矩阵,以0.80为阈值判定环境因子间的共线性。采用MaxEnt模型自带的刀切法(Jackknife)将19个环境变量在模型中按照训练得分进行排序,将贡献度较低且与其他变量间共线性高的环境变量剔除掉。经过筛选,Bio2、Bio6、Bio8、Bio14、Bio16、Bio19的贡献率较高(表1),将其作为最优环境变量纳入后续模型分析。
表1 多花黑麦草生态位分化分析使用的环境因子及其相对贡献值Tab.1 The environmental factors and their relative contribution values used in the niche shift analysis of Lolium multiflorum Lam. |
| 因子类型 | 变量 | 变量描述 | 单位 | 相对贡献值/% |
|---|---|---|---|---|
| Bio2 | 昼夜温差月均值 | ℃ | 1.2 | |
| Bio6 | 最冷月份最低温度 | ℃ | 37.1 | |
| Bio8 | 最湿季度平均温度 | ℃ | 0.9 | |
| 气候因子 | Bio14 | 最干月份降水量 | mm | 23.0 |
| Bio16 | 最湿季度降水量 | mm | 2.4 | |
| Bio19 | 最冷季度降水量 | mm | 35.3 |
1.2.2 生态位模型分析
1.2.3 适生区预测
将多花黑麦草在中国、除中国外的全球分布数据分别和筛选后的环境变量导入MaxEnt软件,采用交叉验证方法运行10次,即随机选取25%的分布点作为测试集,剩余75%作为训练集,设置最大重复次数为5 000。利用刀切法评估各环境变量的相对贡献率,采用受试者工作曲线与横坐标围成的面积值评估模型精度。
1.2.4 入侵阶段分析
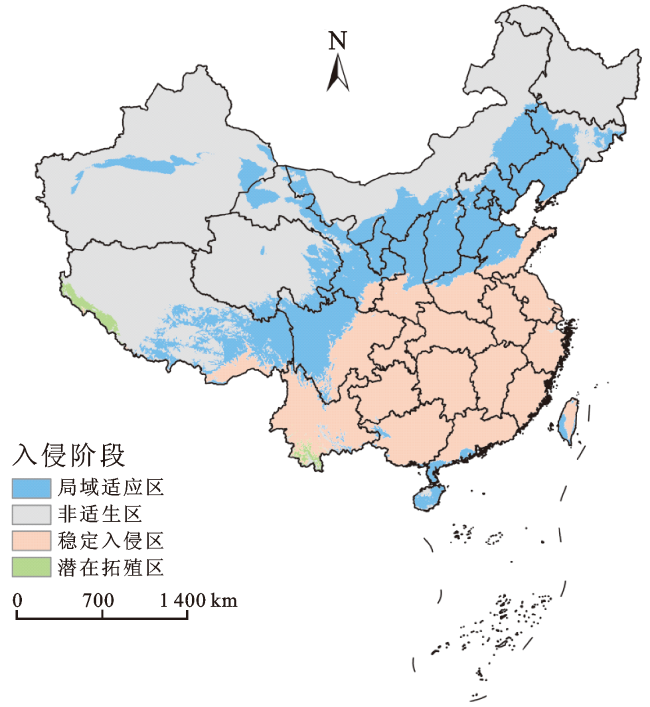
理论上,入侵物种与环境的关系随着入侵阶段的不同而变化。利用Gallien等提出的新框架,可以推断一个物种不同种群在其生态位空间中的入侵阶段,包括局域适应区(位于区域生态位之内,但在全球生态位之外)、非适生区(位于区域生态位和全球生态位之外)、稳定入侵区(位于区域生态位和全球生态位之内)和潜在拓殖区(位于区域生态位之外,但在全球生态位之内),从而能够更可靠地预测外来入侵物种的分布,提高对其入侵风险的评估[24]。
采用最大训练灵敏度加特异性逻辑阈值(maximum training sensitivity plus specificity logistic threshold, MTSS)将基于中国分布点的预测结果与基于除中国外全球区域的预测结果通过赋值转化为0/1二元结果;再将二者的图层进行相减得到多花黑麦草在中国的入侵阶段分布图。基于中国分布点的预测结果的MTSS阈值为0.122 0,基于除中国外全球区域的预测结果的MTSS阈值为0.101 8。
1.2.5 数据分析
生态位模型构建与分析采用ecospat、factoextra、rgdal、sf、adehabitatMA、ade4、raster等程序包在R 4.1.3软件中完成,适生区预测利用MaxEnt 3.3.3完成,模型结果图示化在DIVA-GIS和ArcGIS 10.2软件中完成。本研究所使用的底图数据是基于自然资源部标准地图服务网站(http://bzdt.ch.mnr.gov.cn)下载的中国地图(审图号:GS(2020)4619号)。
2 结果与分析
2.1 生态位分化
主成分分析结果显示,多花黑麦草原产地种群与在中国的入侵地种群的生态位重叠度极低,种群间的生态位相似性极低(D=0.074,P<0.05)。环境因子贡献率的分析结果显示,昼夜温差月均值(Bio2)、最干月份降水量(Bio14)、最湿季度降水量(Bio16)对PC1轴的贡献较大,最湿季度平均温度(Bio8)对PC2轴的贡献较大。PC1轴解释了研究区环境因子方差的47%,PC2轴解释了研究区环境因子方差的21.87%(图2)。
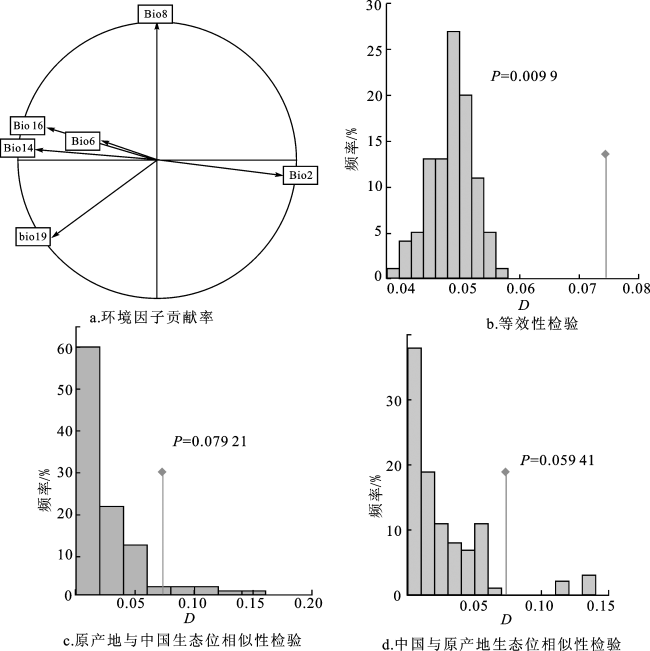
多花黑麦草在入侵中国后PC1轴的生态位宽度明显高于原产地生态位(图3)。与原产地种群相比,多花黑麦草在入侵中国过程中其生态位发生了质心偏移,即生态位漂移(稳定指数仅为0.134,未填充指数为0.848);同时,其生态位也发生显著扩展(扩展指数为0.866)。结果说明多花黑麦草在入侵中国后其生态位发生了显著进化。
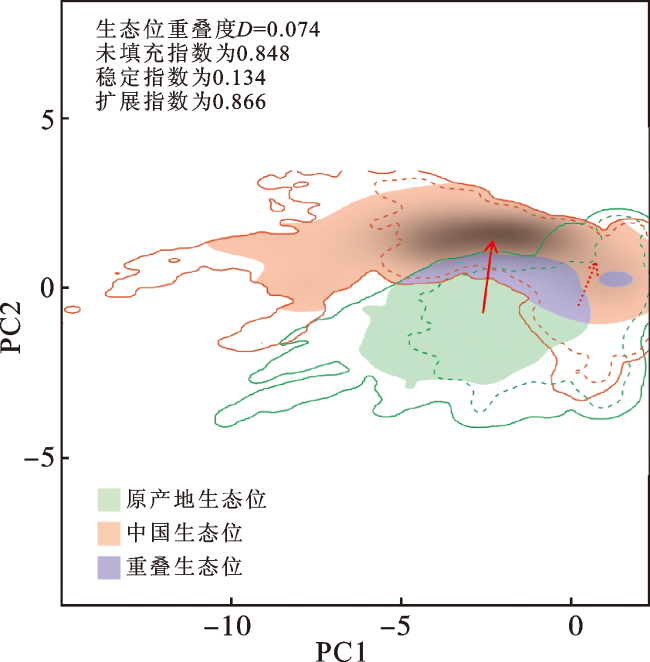
图3 多花黑麦草入侵中国后的生态位分化分析结果注:PC1和PC2表示PCA的前2个轴。实线和虚线分别代表研究区域100%和50%的可用环境。红色箭头显示了多花黑麦草的气候生态位质心(实线)和背景范围(虚线)在2个范围之间的移动情况。生态位动态指数中,0.134为稳定指数(存在于原产地和入侵地生态位空间的比例),0.848为未填充指数(仅存在于原产地范围内的生态位空间的比例),0.866为扩展指数(仅存在于入侵地范围内的生态位空间相对于原产地范围的比例)。网络版为彩图。 Fig.3 The results of niche shift analysis after the invasion of Lolium multiflorum Lam. in China |
2.2 在中国的潜在适生区分布
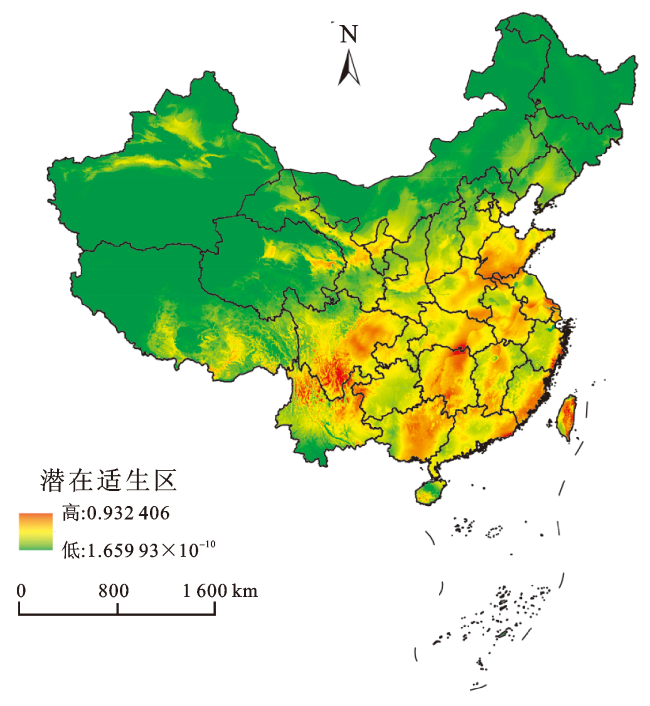
基于多花黑麦草在中国的气候生态位预测结果,可以看出其在我国的中、高度适生区多集中在华中、华南、华东及西南东南部(图4)。其中,云南东北部、四川东南部、福建东南沿海地区、湖北东部、湖南、山东、上海、广西以及台湾等地是多花黑麦草的高度适生区,这些区域面临的入侵风险等级较高。
基于多花黑麦草在除中国外的全球气候生态位预测结果,可以看出多花黑麦草的中、高度适生区多集中在华中和华东地区(图5)。其中,湖南、江西、福建、浙江、上海、贵州东部、广西和广东北部、安徽和湖北南部为多花黑麦草的高度适生区。
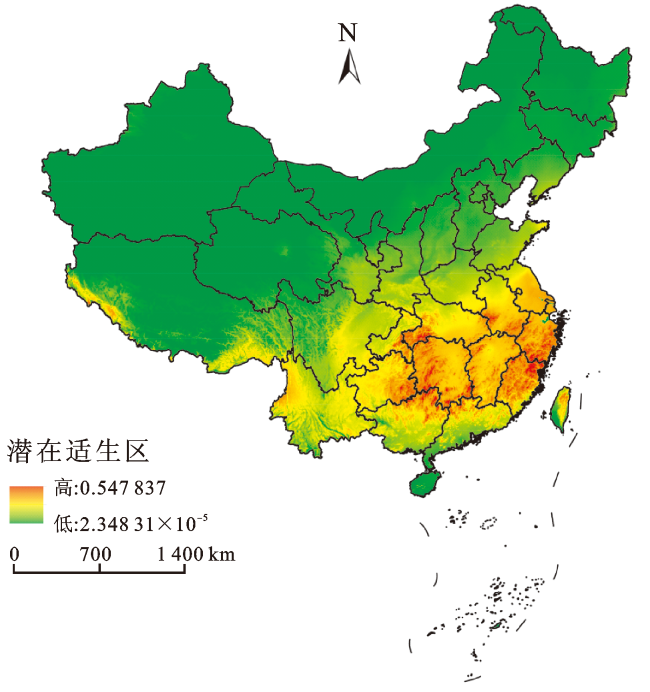
与基于中国种群气候生态位预测的多花黑麦草在中国的潜在适生区分布相比,基于除中国外全球气候生态位所预测的其在中国的潜在适生区分布范围更小、更集中;其中,湖南、江西、福建、浙江、贵州的高度适生区面积明显增加。而基于中国种群气候生态位所预测的其在中国的潜在适生区分布范围比较广,且多花黑麦草有向更北区域扩散的趋势,例如,青海、新疆、内蒙古等地。
2.3 入侵阶段分析
入侵阶段分析结果显示,多花黑麦草在我国的潜在拓殖区(绿色)范围很小,仅在西藏东部、云南南部有极少量分布(图6);稳定入侵区(粉色)范围很广,包括南方大部分省份以及小部分北方地区(山东东南部、陕西南部、甘肃东南部);多花黑麦草在山西、辽宁、北京、天津、河北、宁夏、海南、甘肃南部、陕西北部、山东北部、四川西部、内蒙古东南部、西藏东部等地产生了局域适应性(蓝色),该种在这些区域的入侵正是其生态位进化的结果。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 讨论
生态位保守性是运用生态位模型预测物种分布的前提条件[7]。宋兴江等使用MaxEnt模型研究豚草的生态位时,发现豚草在大多数入侵地区保留了其原产地的气候生态位特征[25],这与生态位保守主义的预期假设一致。然而,本研究发现多花黑麦草在入侵中国的过程中生态位保守性低,导致其适生区预测可能并不准确。有研究表明,使用主成分分析方法对物种在入侵地和原产地的生态位进行量化,并同时使用物种在入侵地和原产地的分布数据在全球尺度进行入侵风险评估,可以在一定程度上消除模拟结果的不确定性[26]。本研究通过环境主成分分析模型对比了多花黑麦草原产地和入侵地种群的生态位差异,为多花黑麦草在入侵中国过程中生态位发生进化提供了证据。与原产地种群相比,多花黑麦草在入侵我国后生态位发生了漂移,同时其生态位也发生显著扩展。在入侵过程中,外来植物气候生态位发生变化的现象普遍存在,这种生态位变化有助于物种入侵新的地区或扩大其入侵地区的分布[27],对入侵物种扩散具有重要影响。
多花黑麦草喜温暖湿润气候,最适在年降水量为1 000~1 500 mm的地区生长[28]。它在我国的入侵趋于向更北的中温带和更南的热带地区扩展,表明其在入侵过程中产生了局域适应,该种在这些区域的入侵是其生态位进化的结果。例如,根据除中国外的多花黑麦草种群生态位预测结果,陕西省关中平原和河南中部等地属于低度适生区;然而,根据中国种群的生态位预测结果,这些地方是该种的中、高度适生区。事实上,根据实地调查结果,多花黑麦草在关中平原和河南省已大规模暴发,并对当地农业生产等造成了较大影响[3]。物种在入侵过程中,其生态位和实际分布可能会受到物种相互作用的显著影响[29];此外,气候变化[30]、人类活动[31]等也可能会使入侵种的适生区范围发生改变。在本研究中,入侵地中国的多花黑麦草生态位质心主要沿着第二轴向上移动。根据主成分分析结果(图2a),第二轴主要反映的是温度变化(Bio8),因此多花黑麦草的局域适应主要体现在相较于原产地种群,其在中国的入侵种群适应了更高温的环境,比如中国雷州半岛、海南、台湾南部等热带地区。相似的研究表明,粗毛牛膝菊(Ganlisoga quadriradiata)在入侵中国的过程中也发生了明显的局域适应,但其适应的环境趋向于比原产地更冷和更干旱的区域[29]。这些研究结果说明外来入侵植物在扩散过程中发生局域适应并导致生态位扩展的现象是普遍存在的。入侵物种到达入侵地后,入侵地的环境选择压力可能与原产地存在较大差异,在选择压力的作用下,入侵种群对环境因子的适应能力会发生改变,尤其是对温度和降水的适应。因此,在对入侵种扩散动态的预测研究中,应该关注局域适应可能带来的影响,仅基于原产地种群生态位预测的入侵种扩散潜力往往会低估入侵种的实际入侵潜力。
本研究结果说明,无论是基于中国种群气候生态位预测的适生区分布模型,还是基于除中国外全球气候生态位所预测的适生区分布模型,多花黑麦草在我国中东部都具有较高的潜在入侵风险,例如湖南、湖北、江西、福建、安徽、上海等地区均为多花黑麦草的中、高度适生区。现有的分布记录也显示(图1),上述地区正是多花黑麦草在我国的主要危害区域,这些地方应高度重视对该种的防控。此外,模型结果也表明,中国仍存在一些未填充区域,多花黑麦草在中国的入侵种群尚未达到区域平衡,仍有向新气候适宜区扩散的可能。一方面,多花黑麦草具有种子量大、繁殖能力强、再生性强等生物学特性,适应性强,适生范围广;另一方面,有研究表明多花黑麦草具有较高的遗传多样性,遗传变异丰富[32],可为其适应新气候提供基础。据记载,多花黑麦草大约于20世纪30年代被引入我国[2],引入历史尚不太长,该物种主要借助粮食运输或其他农业活动实现远距离扩散,因此仍有大面积的潜在适生区域尚未被该种占据。随着人为干扰活动的加强,该物种可能会进一步扩张其分布区,占据尚未被填充的区域。
作为恶性禾本科杂草,多花黑麦草的传播速度非常快。据调查,2015年以来,在陕西、河南、山东、山西、河北等省份麦田发生危害的面积已达86万km2,其中以多花黑麦草为代表的恶性杂草对小麦的危害程度越来越严重,威胁到小麦的生长安全和粮食作物安全[16]。在河南省对麦田杂草的调查中发现,多花黑麦草对花生-小麦连作区危害严重,造成小麦产量损失20%以上[33]。多花黑麦草在陕西的发生面积已从1997年的1.2万km2增加到目前的30万km2以上,涉及西安、渭南、铜川、咸阳等市的近30多个县区,成为陕西省小麦高产稳产的最大屏障[16]。由于多花黑麦草根系发达、分蘖和繁殖能力强,其入侵麦田后与小麦争夺水肥,对小麦生产构成严重威胁,必须采取措施抑制其蔓延和危害。
一方面,严格把控作物种子质量,限制其扩展速度;另一方面,通过科学用药,提高化学防治效果。比如小麦播种后发芽前用砜吡草唑悬浮剂(40%)封闭除草,对已播种完毕且有多花黑麦草的地块可选用5%唑啉草酯混配碇磺草胺、甲基二黄隆等进行防治[18]。此外,还应通过耕地旋耙和人工拔除,清除其根系,消灭麦田中残存的多花黑麦草。当前,多花黑麦草防治主要依赖的还是化学防治方法。这种方法虽然效果较好,但长期大量施用化学药剂可能造成环境污染和农药残留。未来针对多花黑麦草的防控应该在尚未暴发区域采取预防为主的治理策略,而对于多花黑麦草已经暴发的地区可加强推进防治相关研究,比如开发环境友好型的生物农药或者生物菌剂等。
4 结论
综上所述,多花黑麦草入侵我国后生态位发生了显著漂移,其在我国东中部大部分地区具有较高的入侵潜力。本研究有助于构建多花黑麦草入侵风险预警,为开展多花黑麦草的防控和治理提供了重要研究基础。在此基础上,应根据多花黑麦草的潜在分布,对其传播和扩散动态加强监测,加强该物种的跨省跨区域检验检疫,遏制其快速扩散蔓延的势头,防止大面积爆发,降低对我国农业生产和区域生态安全可能造成的危害。