

1 材料与方法
1.1 材料
1.2 培养光强设置
1.3 最大光化学量子产量(Fv/Fm)和非光化学淬灭的测定
1.4 快速叶绿素荧光诱导动力学曲线参数的测定
1.5 快速光响应曲线的测定
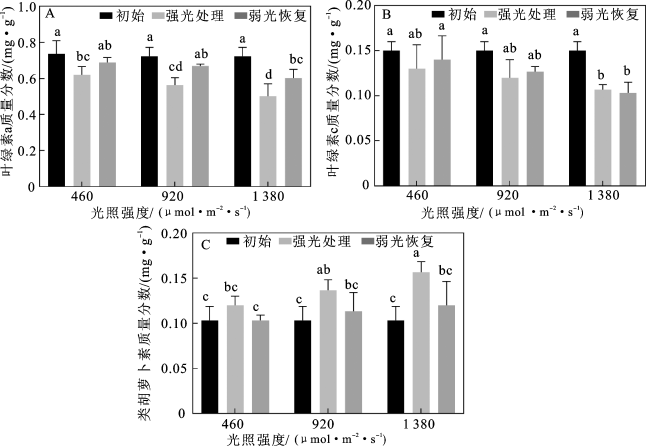
1.6 光合色素含量的测定
1.7 数据分析
2 结果
2.1 叶绿素荧光参数
表1 不同时间和光强条件下Fv/Fm和qN的双因素重复测量方差分析结果Tab.1 Results of RM-ANOVA for Fv/Fm and qN under different time and light intensity conditions |
| 因 素 | Fv/Fm | qN | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | P | df | F | P | |||||||
| 时间 | 8 | 83.454 | <0.001 | 8 | 31.230 | <0.001 | ||||||
| 光强 | 2 | 38.825 | <0.001 | 2 | 12.349 | 0.007 | ||||||
| 时间×光强 | 16 | 3.719 | <0.001 | 16 | 0.737 | 0.743 | ||||||
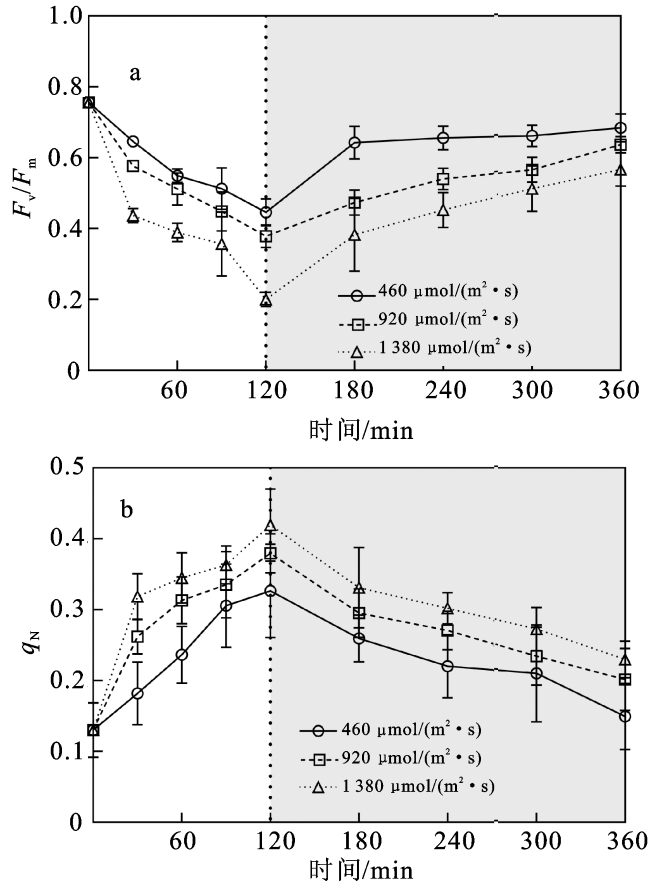
2.2 快速叶绿素荧光诱导动力学曲线参数
表2 不同时间和光强条件下VJ、M0和ψ0的双因素重复测量方差分析结果Tab.2 Results of RM-ANOVA for VJ,M0,ψ0 under different time and light intensity conditions |
| 因 素 | VJ | M0 | ψ0 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | P | df | F | P | df | F | P | ||||||||||
| 时间 | 8 | 31.321 | <0.001 | 8 | 55.232 | <0.001 | 8 | 31.712 | <0.001 | |||||||||
| 光强 | 2 | 25.968 | 0.001 | 2 | 233.498 | <0.001 | 2 | 24.924 | 0.001 | |||||||||
| 时间×光强 | 16 | 1.195 | 0.306 | 16 | 5.653 | <0.001 | 16 | 1.456 | 0.157 | |||||||||
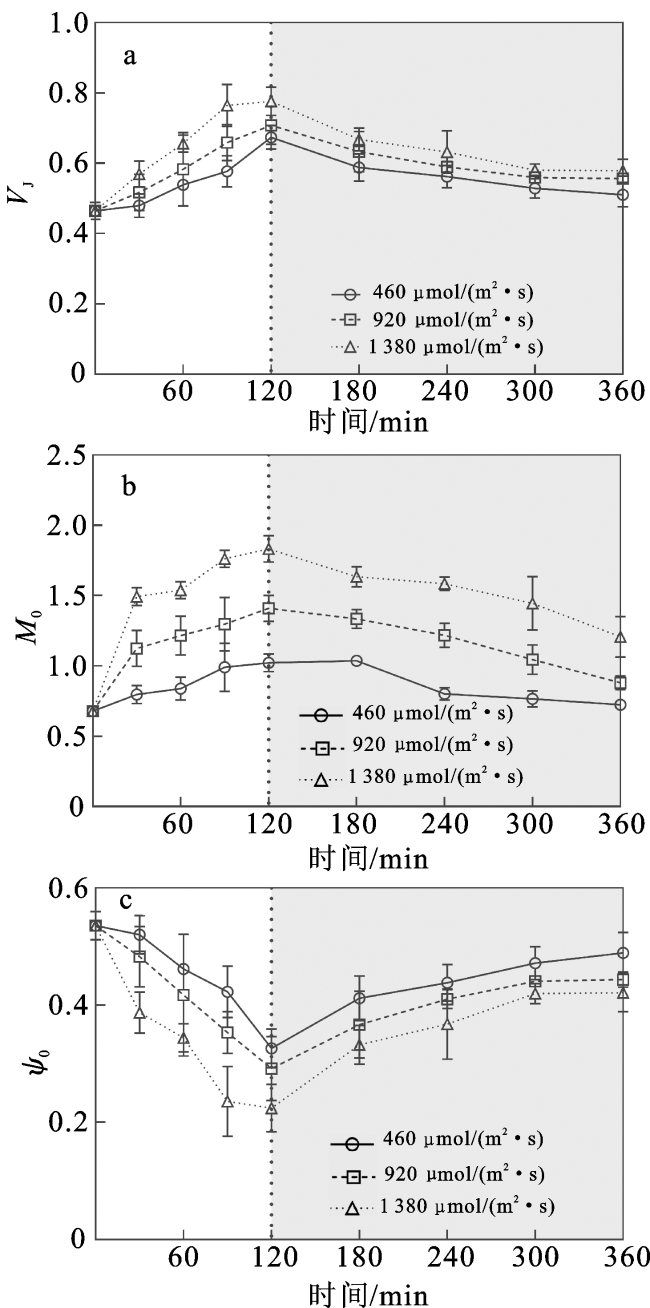
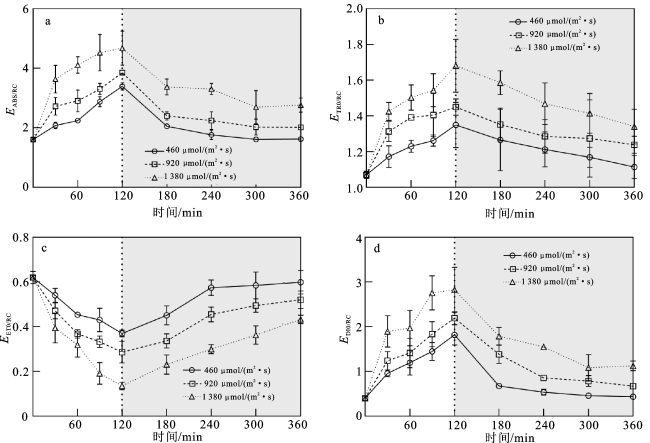
图3 强光处理(白色背景部分)和弱光恢复(灰色背景部分)过程中EABS/RC(a)、 (b)、 (c)、 (d)的变化注:120 min时3种强光处理的藻样均转到相同弱光条件下进行恢复培养。 Fig.3 Changes in EABS/RC (a), (b), (c), (d) of Sargassum horneri during the strong light treatment(white background) and the weak light recovery (grey background) process |
表3 不同时间和光强条件下EABS/RC、ETR0/RC、EET0/RC和EDI0/RC的双因素重复测量方差分析结果Tab.3 Results of RM-ANOVA for EABS/RC,ETR0/RC,EET0/RC,EDI0/RC under different time and light intensity conditions |
| 因 素 | EABS/RC | ETR0/RC | EET0/RC | EDI0/RC | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | P | df | F | P | df | F | P | df | F | P | |||||||||||||
| 时间 | 8 | 61.772 | <0.001 | 8 | 20.459 | <0.001 | 8 | 73.794 | <0.001 | 8 | 72.222 | <0.001 | ||||||||||||
| 光强 | 2 | 439.305 | <0.001 | 2 | 15.445 | 0.004 | 2 | 204.686 | <0.001 | 2 | 74.919 | <0.001 | ||||||||||||
| 时间×光强 | 16 | 2.842 | 0.003 | 16 | 1.307 | 0.232 | 16 | 3.560 | <0.001 | 16 | 2.644 | 0.005 | ||||||||||||
2.3 快速光响应曲线
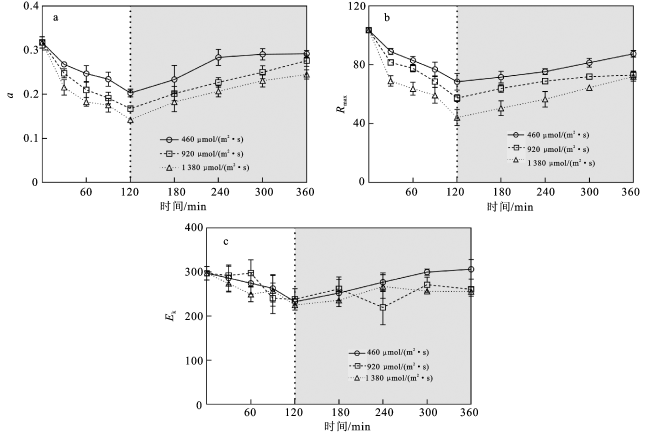
图4 强光处理(白色背景部分)和弱光恢复(灰色背景部分)过程中铜藻快速光响应曲线的α(a)、Rmax(b)、Ek (c)的变化注:120 min时3种强光处理的藻样均转到相同弱光条件下进行恢复培养。 Fig.4 Changes in α(a),Rmax(b) and Ek (c) of the rapid light response curve during the strong light treatment (white background) and the weak light recovery (grey background) process |
表4 不同时间和光强条件下α、Rmax和Ek的双因素重复测量方差分析结果Tab.4 Results of RM-ANOVA for α,Rmax and Ek under different time and light intensity conditions |
| 因 素 | α | Rmax | Ek | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | P | df | F | P | df | F | P | |||||||||||
| 时间 | 8 | 86.247 | <0.001 | 8 | 267.223 | <0.001 | 8 | 6.885 | <0.001 | ||||||||||
| 光强 | 2 | 58.106 | <0.001 | 2 | 38.227 | <0.001 | 2 | 7.583 | 0.023 | ||||||||||
| 时间×光强 | 16 | 2.089 | 0.025 | 16 | 7.579 | <0.001 | 16 | 1.691 | 0.081 | ||||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}