

植物膜受体蛋白能够感知外界刺激,识别多种胞外及胞间信号分子,在植物激素信号传导、生长发育、非生物胁迫防御和抗病等生理过程中起重要作用。富含亮氨酸重复受体蛋白(leucine-rich repeat receptor like proteins, LRR-RLPs)是植物膜蛋白家族中的一种,其成员具有胞外LRR结构域,但缺乏胞内激酶结构域[1-2]。胞外LRR结构域是RLPs蛋白最常见的配体结合区域,能识别多种受体,包括固醇、脂质、糖、肽、脂肽和核酸等[3]。此外,胞外LRR结构域通常含有许多潜在的糖基化位点,参与胞内信号传导[2,4]。LRR-RLPs蛋白广泛存在于高等植物中,目前已在部分植物基因组中鉴定出LRR-RLPs家族成员,包括拟南芥(57个)[1]、水稻(90个)[5]、番茄(176个)[6]、普通烟草(70个)[7]、油菜(276个)[8]和藜麦(106个)[9]等。
前期研究表明,多数LRR-RLPs蛋白参与调控植物的抗病和抗逆等生理过程。例如,Cf-9和Cf-4能激活番茄对褐孢霉的抗性基因[10-11],Ve1和Ve2介导番茄对黄萎病的抗性[12-13],AtRLP1、AtRLP3、AtRLP23、AtRLP30和AtRLP42调控拟南芥对多种病原菌的识别和免疫反应激活[14-18],水稻OsRLP1蛋白与其适配激酶OsSOBIR1互作能激活寄主对病毒感染的免疫[19],AtLRRop2和COG1基因分别参与拟南芥和水稻的低温或冷冻胁迫反应[4,20]。部分LRR-RLPs蛋白调控模式植物拟南芥和水稻生长发育的作用也已被证实。例如,AtRLP17/TMM、AtRLP10/CLV2和AtRLP44分别参与拟南芥的气孔发育、顶端分生组织增殖和细胞壁发育[21-23],AtRLP41调控拟南芥叶片对脱落酸(abscisic acid, ABA)的敏感性[1],YPD1蛋白参与水稻叶绿体的发育和强光诱导的叶片衰老[24]。然而,RLPs蛋白调控荞麦属(Fagopyrum Mill)植物生长发育的研究尚未见报道。
苦荞(Fagopyrum tataricum),又名鞑靼荞麦(Tartary buckwheat),是荞麦属植物的两个栽培种之一。苦荞作为我国重要的药食两用杂粮作物,其籽粒不仅富含膳食纤维、脂肪和维生素等营养物质,还富含黄酮、多酚等功能化合物,具有显著的抗氧化、抗炎、降三高(高血脂、高血压和高血糖)及抗癌作用[25-27],在食品、医药、保健和饲料等领域均具有很大发展潜力。中国的苦荞种植面积和产量居全球首位,但亩产低(170~250 kg/hm2)[28]且品质参差不齐,在国际上仍处于供不应求的状态,因此提高苦荞产量和品质是发展苦荞产业亟待解决的问题。苦荞千粒重主要由粒形(粒长、粒宽、长宽比)和充实度决定,是影响苦荞产量、外观和营养品质的重要农艺性状[29-32]。研究表明,苦荞千粒重具有较高遗传力[33-34],不易受环境因素的影响,是提高苦荞产量和品质的最稳定因素。因此,挖掘和利用苦荞粒重和粒形调控基因越来越受到苦荞育种工作者的重视。
近年来,大宗作物粒形和粒重相关的数量性状位点(quantitative trait,loci,QTL)定位、基因克隆及功能分析取得了一定的研究进展[35]。与大宗作物相比,苦荞粒重和粒形性状的分子研究还处于起步阶段。目前,已有利用高通量测序技术鉴定苦荞粒重和粒形性状QTL的报道[34,36 -38],但关键候选基因的功能还未验证。本课题组前期定位到1个同时控制千粒重和粒形的主效QTL,并筛选到1个关键候选基因FtPinG00001423200.01,其既在双亲间存在SNP/InDel变异,也在双亲籽粒不同发育时期存在表达差异,该基因与拟南芥RLP48蛋白同源性最高,故命名为FtRLP48,推测其可能参与苦荞籽粒发育的调控[34]。为验证该基因的功能,本研究在初级定位群体双亲‘小米荞’和‘晋荞麦2号’中克隆得到该基因,验证其SNP/InDel变异,并对其进行生物信息学和亚细胞定位分析,通过转基因技术明确其对籽粒发育的影响,为苦荞粒重形成的分子调控机理研究以及分子标记辅助选育优异苦荞品种提供理论依据。
1 材料与方法
1.1 实验材料
供试植物材料为厚壳、长粒、大粒品种‘晋荞麦2号’和薄壳、短粒、小粒品种‘小米荞’,种植于贵州师范大学荞麦工程技术研究中心安顺基地。质粒pMD19-T购自宝日医生物技术(北京)有限公司。大肠杆菌(Escherichia coil)DH5α感受态细胞、根瘤农杆菌(Agrobacterium tumefaciens)GV3101感受态细胞、质粒pBI121、水稻品种‘中花11’和本氏烟草(Nicotiana benthamiana(K.))由本实验室保存。
1.2 总RNA提取及第一链cDNA合成
用镊子采取两亲本授粉后15 d的籽粒,立即放入液氮速冻,利用E.Z.N.A Plant RNA Kit试剂盒(Omega)提取总RNA。采用FastKing cDNA第一链合成试剂盒(北京天根生化科技有限公司)反转录,合成用于基因克隆的第一链cDNA。
1.3 全长CDS克隆
根据苦荞参考基因组中FtRLP48的CDS序列(FtPinG00001423200.01)[39],利用Primer 5.0软件设计扩增FtRLP48的全长特异性引物(表1),利用Primer STAR MAX DNA Polymerasa高保真酶(TaKaRa)以1.2中的cDNA为模板进行PCR扩增。反应程序为98 ℃ 3 min;98 ℃ 10 s,56.8 ℃ 15 s,72 ℃ 2 min,35个循环;72 ℃ 10 min;4 ℃ 30 min。用1%琼脂糖凝胶电泳检测扩增产物,切下片段大小正确的目标条带,用FastPure Gel DNA Extraction Mini Kit试剂盒(Vazyme)进行胶回收,Tap酶(TaKaRa)进行加A尾反应后,连接pMD19-T载体(TaKaRa)。随后转化到大肠杆菌感受态细胞DH5α中,涂在含有氨苄青霉素抗生素的LB培养基平板上,37 ℃过夜培养后,挑选单克隆进行PCR检测,送阳性克隆至生工生物工程(上海)股份有限公司测序。将‘晋荞麦2号’和‘小米荞’为模板构建的重组质粒分别命名为pMD19-T-FtRLP48-J和pMD19-T-FtRLP48-M。
表1 本研究所用引物序列Tab.1 Primer sequences used in this study |
| 引物名称 | 引物序列(5'-3') | 用途 |
|---|---|---|
| FtRLP48 | F:ATGTTGGTCTTGATCATCCAATTACT R:TTATTTATACAATCTTATTGAGATTTTTTTCA | 基因克隆 |
| pBI121-FtRLP48 | F:ggactctagaggatccccgggATGTTGGTCTTGATCATCCAATTACT R:cgatcggggaaattcgagctcTTATTTATACAATCTTATTGAGATTTTTTTCA | 遗传转化 |
| FtRLP48-GFP | F:agagaacacgggggactttgcaacATGTTGGTCTTGATCATCCAATTAC R:cagctcctcgcccttgctcaccatAGATCCTCCTCCAGATCCTCCTCCcgtacatgtatggggtcgac | 亚细胞定位 |
| pBI121(35S) | F:GACGCACAATCCCACTATCC | 转基因水稻阳性株系鉴定 |
| pBI121-FtRLP48-R2 | R:GGTGACATGGCCTGTTGAAG |
注:小写核酸序列为载体的同源臂引物。 |
1.4 生物信息学分析
运用Expasy(https:∥web.expasy.org/protparam/)在线分析网站进行FtRLP48蛋白理化性质分析。利用ProtScale(https:∥web.expasy.org/protscale/)预测蛋白质序列亲水性。使用在线软件SMART-MODEL(http:∥smart.embl-heidelberg.de/)、PRABI-SOPMA(https:∥npsa-pbil.ibcp.fr/cgi-bin/npsa-automat.pl?page=npsa-sopma.html/)和SWISS-MODEL(https:∥swissmodel.expasy.org/)预测蛋白质的结构域、二级结构和三级结构。通过UniProtKB(https:∥www.uniprot.org/blast/)数据库进行FtRLP48同源比对,从比对结果中筛选出evalue值最小且来源于拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、番茄(Solanum lycopersicum)、大豆(Glycine max)、玉米(Zea mays)、黄瓜(Cucumis sativus)、烟草(Nicotiana tabacum)、葡萄(Vitis vinifera)、银白杨(Populus alba)和小麦(Triticum aestivum)注释为LRR-RLPs的蛋白序列,利用T-coffee(https:∥www.ebi.ac.uk/jdispatcher/msa/tcoffee?stype=protein/)在线工具构建系统发育树,然后将结果以Newick格式导入iTOL(https:∥itol.embl.de/tree/)在线工具绘制系统发育树。
1.5 FtRLP48蛋白的亚细胞定位
1.6 水稻遗传转化
采用同源重组的方法将‘晋荞麦2号’和‘小米荞’FtRLP48的CDS序列分别连接到过表达载体pBI121上,构建过表达载体pBI121-FtRLP48-J和pBI121-FtRLP48-M,过表达载体的构建参照赵鑫等[41]的方法进行。将2个重组质粒转化至农杆菌EHA105感受态细胞中,筛选鉴定得到阳性克隆,将阳性菌液扩培至吸光度OD600到0.2。将种子进行机械脱壳,用75%酒精消毒,无菌水清洗干净后,播种于愈伤组织诱导培养基上,26 ℃光培养20 d。水稻愈伤组织诱导、遗传转化和培养的具体操作参考唐彬[42]的方法进行。采用CTAB法提取水稻基因组DNA作为模板,以pBI121载体通用引物pBI121(35S)为正向引物,以在FtRLP48-J和FtRLP48-M一致序列5'端设计的基因特异性引物pBI121-FtRLP48-R2为反向引物进行PCR反应(表1),经检测后获得pBI121-FtRLP48-J和pBI121-FtRLP48-M单株阳性过表达植株,挑选阳性苗和野生型‘中花11’水稻(WT)进行种植。
1.7 千粒重和粒形性状的调查
收取转基因和WT水稻成熟单株,脱粒,选取饱满的种子约100粒,利用万深SC-G型自动种子考种仪测定千粒重、粒长、粒宽和长宽比等性状;选取饱满的种子50粒,手动剥脱壳后,记录籽粒果壳重和果仁重。采用IBM SPSS Statistics 26软件对各性状进行描述统计分析、方差分析和多重比较(Ducan法)。
2 结果与分析
2.1 FtRLP48基因克隆及编码蛋白特性分析
利用基因特异引物FtRLP48-F和FtRLP48-R,以授粉15 d后的籽粒cDNA为模板进行PCR扩增,扩增目的条带约2 500 bp(图1),将在‘小米荞’和‘晋荞麦2号’中克隆得到的基因分别命名为FtRLP48-M和FtRLP48-J。测序和比对结果显示:FtRLP48-M与参考基因组中该基因的序列一致,基因大小为2 577 bp,编码858个氨基酸;FtRLP48-J的扩增条带大小为2 573 bp,其在1 902 bp处的“ATGGA”变异为“C”,导致FtRLP48-J在1 917 bp处提前形成1个终止密码子 TAA, 翻译提前终止,共编码638个氨基酸,比FtRLP48-M少编码220个氨基酸(图2、图3)。
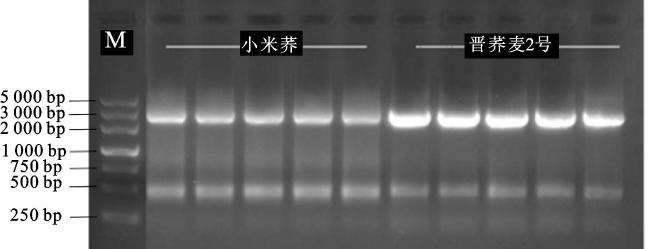

图2 FtRLP48在‘小米荞’和‘晋荞麦2号’中碱基序列比对注:网络版为彩图。 Fig.2 Base sequence alignment of FtRLP48 in ‘Xiaomiqiao’ and ‘Jinqiaomai 2’ |

2.2 FtRLP48的生物信息学分析
根据参考基因组提供的注释信息,分析FtRLP48的基因结构,结果显示,该基因不含内含子,其起始密码子为ATG,终止密码子为TAA。将FtRLP48蛋白序列在UniProtKB数据库进行BLASTp比对,发现该蛋白序列与拟南芥AtRLP48(F4JTU7)的蛋白序列相似性最高(35.2%)。
2.2.1 FtRLP48蛋白理化性质分析
Expasy在线软件分析结果表明:FtRLP48-J和FtRLP48-M蛋白的分子式分别为C4316H6802N1130O1264S29和C3165H5016N838O940S22,相对分子量分别为95.68 kDa和70.55 kDa,等电点分别为6.22和6.19。ProtScale在线软件分析显示:FtRLP48-J和FtRLP48-M蛋白的平均亲水性分别为0.056和0.067,不稳定系数分别为38.10和40.52,推测FtRLP48-M是亲水性的稳定蛋白,而FtRLP48-J是亲水性的不稳定蛋白。
2.2.2 FtRLP48蛋白二级结构和三级结构预测
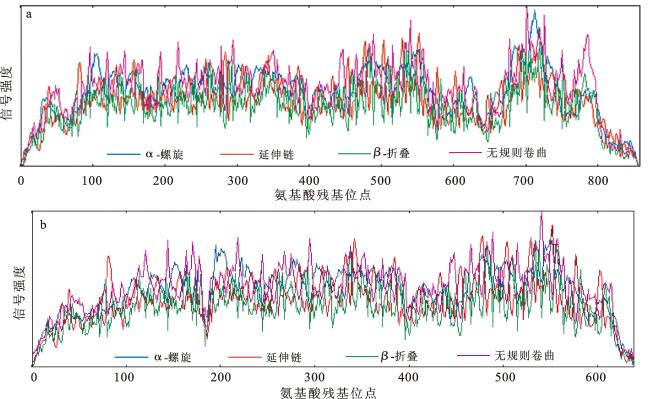
图4 FtRLP48-M(a)和FtRLP48-J(b)蛋白的二级结构预测注:网络版为彩图。 Fig.4 Secondary structure prediction of FtRLP48-M (a) and FtRLP48-J (b) proteins |
2.2.3 FtRLP48蛋白结构域分析
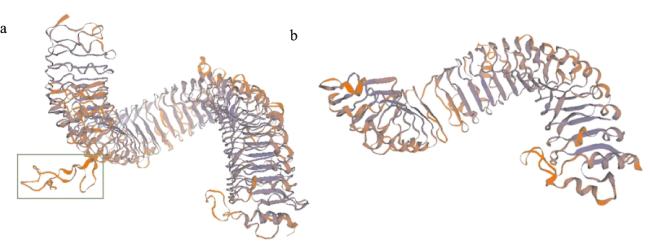
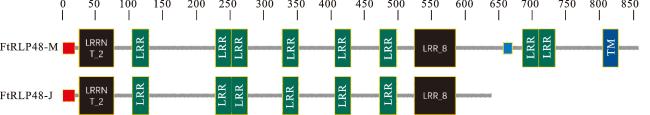
SMART-MODEL分析结果显示,FtRLP48-M蛋白有N端信号肽、富含亮氨酸的重复N端结构域(LRRNT-2)、富含亮氨酸重复域(LRR)、跨膜结构域(TM)和不具备激酶结构域的细胞外短肽,具有RLPs基因家族编码蛋白的典型特征,属受体类蛋白;而FtRLP48-J蛋白缺失了跨膜结构域和细胞外短肽(图6)。
2.2.4 FtRLP48的系统进化树分析
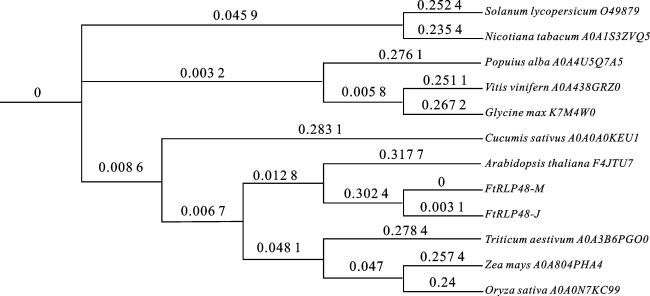
比较FtRLP48与其他物种RLP的亲缘关系,利用FtRLP48蛋白序列在UniProtKB数据库进行BLASTp比对,获得拟南芥、水稻、玉米、番茄和大豆等其他10个物种的RLP蛋白序列,与FtRLP48-M和FtRLP48-J的蛋白序列进行比对,构建系统进化树。如图7所示, FtRLP48-M和FtRLP48-J的蛋白序列相似度最高,二者与拟南芥在进化树上遗传距离最近,其次是与单子叶粮食作物小麦、玉米和水稻的亲缘关系较近。
2.3 FtRLP48蛋白的亚细胞定位分析
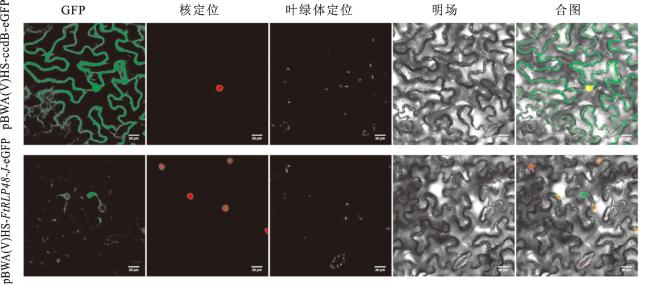
将构建的pBWA(V)HS-FtRLP48-J-eGFP融合蛋白载体和pBWA(V)HS-ccdB-eGFP(对照)通过农杆菌介导法注射侵染本氏烟草叶片,进行瞬时表达,以核定位载体pBWA(V)HS-FtRLP48-J-eGFP为内参,在共聚焦显微镜下观察GFP的荧光信号。如图8所示,对照组空载体的荧光信号分布在烟草表皮细胞的细胞膜和细胞核上,而pBWA(V)HS-FtRLP48-J-eGFP融合蛋白的荧光信号分布在部分细胞膜和细胞核核膜上,由此可得,FtRLP48-J蛋白定位在细胞膜和细胞核核膜上。
2.4 FtRLP48转基因水稻阳性株系鉴定
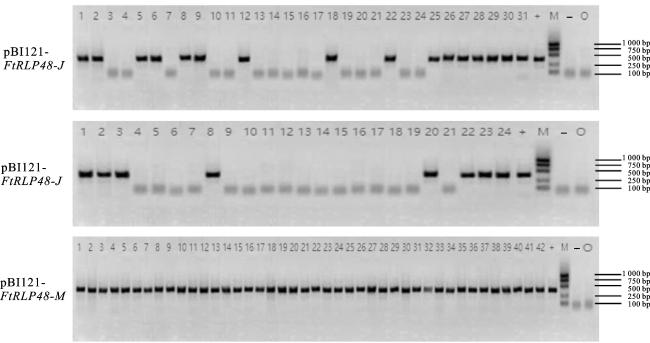
分别构建FtRLP48-M和FtRLP48-J的植物过表达载体,进行水稻遗传转化。以野生型水稻‘中花11’(WT)和T0代转基因水稻植株的DNA为模板,以pBI121载体通用引物pBI121(35S)为正向引物,基因特异性引物pBI121-FtRLP48-R2为反向引物进行PCR验证,结果显示(图9),WT和空白对照未扩增出目的条带,pBI121-FtRLP48转基因植株的PCR扩增产物条带大小与阳性对照条带基本一致,后期分别得到21株pBI121-FtRLP48-J和37株pBI121-FtRLP48-M转基因水稻植株。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
2.5 FtRLP48转基因植株的粒重和粒形分析
FtRLP48-J、FtRLP48-M转基因植株及WT植株的籽粒性状调查结果显示,pBI121-FtRLP48-J转基因水稻植株的粒长和粒宽显著高于WT,分别提高3.27%和3.87%;其千粒重、千粒果壳重和千粒果仁重极显著高于WT,分别增加18.22%、10.49%和16.57%;其长宽比和果壳率与WT没有显著差异。pBI121-FtRLP48-M转基因水稻植株的粒宽和千粒果壳重显著高于WT,分别提高3.55%和8.93%;果壳率极显著高于WT,增加8.51%;长宽比极显著低于WT,降低3.35%;而粒长、千粒重和千粒果仁重与WT没有显著差异(表2)。这说明过表达FtRLP48-J能明显增大粒长和粒宽,极显著提高千粒重,而过表达FtRLP48-M虽能够明显增加粒宽,显著提高果壳重,但对千粒重和果仁重的影响不明显。
表2 野生型及转基因水稻的千粒重和粒形分析Tab.2 Thousand-grain weight and grain shape analysis of WT and transgenic rice |
| 株系 | 植株数 | 粒长/mm | 粒宽/mm | 长宽比 | 千粒重/g | 千粒果壳重/g | 千粒果仁重/g | 果壳率/% |
|---|---|---|---|---|---|---|---|---|
| WT | 3 | 7.35±0.21Ab | 3.10±0.11Ab | 2.39±0.01Aa | 23.32±0.55Bb | 4.48±0.10Bb | 20.70±0.15Bb | 17.75±0.38Bb |
| pBI121-FtRLP48-M | 37 | 7.37±0.16Ab | 3.21±0.09Aa | 2.31±0.06Bb | 23.37±1.48Bb | 4.88±0.25ABa | 20.33±1.18Bb | 19.26±0.93Aa |
| pBI121-FtRLP48-J | 21 | 7.59±0.19Aa | 3.22±0.09Aa | 2.37±0.05ABa | 27.57±1.98Aa | 4.95±0.38Aa | 24.13±1.55Aa | 16.96±1.13Bb |
注:同列数据后不同小写字母表示差异达0.05显著水平,不同大写字母表示差异达0.01显著水平。 |
3 讨论
大粒种子不仅能提高作物产量,还能为植株早期生长提供更多养分,并对非生物胁迫具有一定的耐受性。在模式植物拟南芥和水稻中存在多种信号通路调节种子大小,如蛋白泛素化降解途径、G蛋白信号途径、IKU途径、丝裂原激活途径、植物激素途径、转录因子途径等[35]。目前,苦荞粒重和粒形分子调控机理的研究相对滞后,定位到粒重和粒形性状QTL的数量极少[34,37 -38],相关候选基因的克隆和作用功能还未见报道,极大地限制了苦荞粒重分子调控机制的解析和遗传改良。富含亮氨酸重复受体蛋白(LRR-RLPs)是一类植物最常见的膜受体蛋白,广泛参与植物激素信号传导、生长发育和抗病等过程,但与植物籽粒发育的关系尚未报道。FtRLP48基因是从本课题组前期定位到的千粒重和粒形稳定主效QTL区间内筛选到的1个LRR-RLPs基因家族成员,其在双亲‘小米荞’和‘晋荞麦2号’中存在编码序列变异和籽粒发育时期的表达差异[34],暗示其可能参与调控苦荞籽粒发育。本研究在‘小米荞’和‘晋荞麦2号’中克隆该基因,分别命名为FtRLP48-M和FtRLP48-J。CDS克隆和比对结果显示,FtRLP48-M和FtRLP48-J的编码序列分别为2 577 bp和1 917 bp,分别编码858和638个氨基酸。FtRLP48-J在1 902 bp处发生“ATGGA/C”移码变异,导致其编码区在1 917 bp提前终止。基因结构显示该基因不含内含子。拟南芥57个RLP家族成员中,37个基因(65%)含有单外显子[1]。普通烟草70个RLP家族成员中,45个基因(64.3%)没有内含子。无内含子基因在转录过程中无需经历内含子剪切步骤,是一类响应外界因素快速应答基因,FtRLP48作为一种无内含子基因,可能参与苦荞籽粒发育的调控。
虽然RLP成员之间蛋白序列的整体相似性较低,但都普遍含有3个结构域,分别是胞外富含亮氨酸重复序列、跨膜结构域及胞内的短肽而不是激酶结构域[1]。跨膜结构域在胞内信号分子的识别与信号传导过程中起重要作用,然而一些没有跨膜结构域的LRR-RLPs能以糖基磷脂酰肌醇锚点附着在细胞外表面[43]。本研究中FtRLP48-J翻译提前终止造成跨膜结构和胞内短肽缺失。FtRLP48-J蛋白亚细胞定位于细胞膜和细胞核核膜上,这与大部分RLP蛋白仅定位到细胞膜不符[1,44]。在RLP家族中,部分基因的功能已被验证。例如,在拟南芥中CLV2基因编码RLP蛋白可与CLV3和CLEs结合进行信号传递,调控分生组织的维持和器官生长发育[45];在玉米中发现的fear2基因与CLV2具有许多共同结构特征,fear2基因编码的RLP蛋白可调节花序膨大和花分生组织发育[46]。本研究过表达FtRLP48-J转基因水稻植株的粒长和粒宽均显著高于野生型,并极显著提高了千粒重、千粒果仁重和千粒果壳重;而过表达FtRLP48-M转基因水稻植株仅粒宽、千粒果壳重和果壳率显著高于野生型。以上研究结果表明FtRLP48蛋白跨膜结构域的缺失可能影响了胞内外细胞信号的传递和物质运输,从而导致了其调控籽粒发育的功能差异。FtRLP48可能与CLV2和fear2基因的作用机制相似,通过调控分生组织的增殖和发育来控制籽粒大小,但其进行细胞信号传递并对籽粒发育的细胞学形态调控还有待进一步确定。
4 结论
苦荞FtRLP48-M和FtRLP48-J的CDS全长分别为2 577 bp和1 917 bp。FtRLP48-J在1 902 bp处发生了“ATGGA/C”移码变异,导致蛋白翻译提前终止,造成跨膜结构缺失。过表达FtRLP48-J极显著提升了转基因水稻的千粒重,而过表达FtRLP48-M显著增加了转基因水稻的果壳重。研究结果说明FtRLP48蛋白跨膜结构域的缺失对千粒重起正向调控作用,为今后从分子水平阐明苦荞籽粒发育调控的遗传基础以及为苦荞分子标记辅助育种提供了理论依据及材料支持。