

转录因子(transcription factors,TFs)也称为反式作用因子,是一类特殊的蛋白质,能够调控基因转录过程[1]。它们通过结合DNA上的特定序列(通常是启动子或增强子区域)影响RNA聚合酶的招募及活性,进而调控目标基因的表达水平和模式[2]。转录因子根据其分子结构特征可分为多个家族,如WRKY、MYB、bHLH、NAC、bZIP等。其中,WRKY转录因子是植物中规模最大、功能最多样化的转录因子家族之一[3],广泛参与植物生长发育和对环境变化的响应[4]。WRKY转录因子通常与靶基因启动子中的W-box互作,激活或抑制下游基因的表达,从而在多种生理过程中发挥关键作用[5]。
植物次生代谢物(plant secondary metabolites,PSMs)是植物在生长过程中通过次生代谢途径合成的一类具有生理活性的有机化合物[6]。与植物生长必需的基本成分如氨基酸和碳水化合物(即初级代谢物)不同,次生代谢物是由初级代谢物经过修饰(如甲基化、羟基化、糖基化)形成的,通常并不是植物生存所必需的[7]。PSMs在植物防御系统中起着至关重要的作用,保护植物免受生物和非生物胁迫,如感染、物理伤害、紫外线照射和臭氧侵害等,帮助植物适应不断变化的环境[8]。此外,PSMs还能抵御食草动物的侵害,并在化学、制药、化妆品和香料行业中应用广泛[9];某些PSMs还具有抗癌、抗氧化、抗病毒、抗炎等活性[10]。PSMs主要分为三大类:萜类、酚类及含氮含硫化合物,转录因子在PSMs合成中具有关键调控作用[11]。
WRKY转录因子家族在植物中的功能多样性已被广泛研究,尤其是在调节植物生长发育方面,但WRKY转录因子家族如何精确调控植物次生代谢物的生物合成仍有待深入研究。因此,本综述旨在综合现有的研究成果,阐明WRKY转录因子对植物次生代谢物调控的影响,提供WRKY转录因子在次生代谢物合成调控中的作用概览,为植物次生代谢物代谢工程提供可能的应用方向。
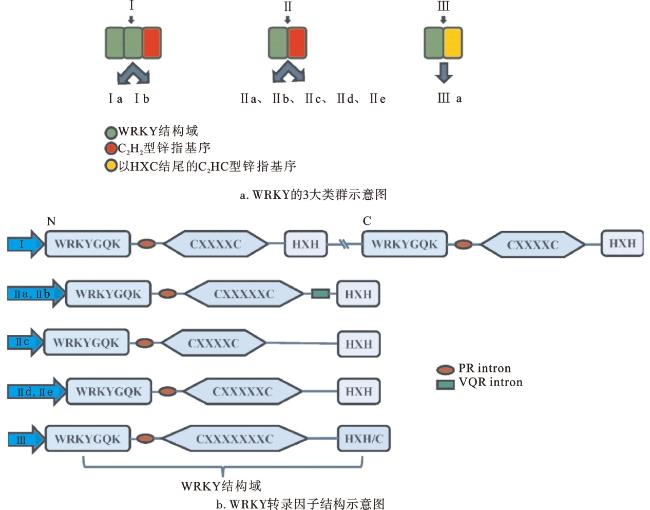
1 WRKY转录因子的结构与分类
WRKY转录因子是调控植物生长发育和响应环境变化的关键因子[12]。WRKY蛋白以1个或2个高度保守的WRKY结构域为特征,大多数WRKY蛋白至少包含1个由约60个氨基酸残基组成的WRKY结构域[13]。WRKY结构域包含2个重要序列,即N端高度保守的七肽序列WRKYGQK(存在变体如WRKYGKK、WRKYGEK等)和C端的金属螯合锌指基序[14-15]。 WRKYGQK序列负责与W-box序列结合,W-box是WRKY转录因子与靶基因启动子特异性结合的重要顺式作用元件,主要存在于抗病虫害、抗旱、抗低温、抗盐碱等相关抗性基因的启动子区域,W-box的核心序列(TGAC/T)是WRKY转录因子结合所必需的,因此WRKY转录因子在植物应对生物和非生物胁迫方面发挥着重要作用[16-17]。C端锌指基序通常为C2H2(CX4-5CX22-23HX1H)型或C2HC(CX7CX23HXC)型,这些结构在WRKY转录因子与DNA的结合过程中起着稳定结构的作用,并有助于增强WRKY转录因子与DNA的亲和力,对WRKY转录因子的功能发挥至关重要[18-20]。在植物进化过程中及不同植物之间,WRKY基因的数量、内含子数目和核苷酸序列都表现出显著差异[2,21]。
根据WRKY结构域的数量和锌指结构的类型,WRKY转录因子可分为3个主要类群(Ⅰ、Ⅱ、Ⅲ,图1a),并可进一步细分为8个亚类[22-23]。类群Ⅰ包含2个WRKY结构域和1个C2H2型锌指基序,主要由C端的WRKY结构域参与DNA结合,而N端WRKY结构域的功能和作用还未得到充分证实。值得注意的是,类群Ⅰ的2个WRKY结构域在功能上可能存在冗余,且其C端WRKY结构域与类群Ⅱ、Ⅲ的WRKY结构域在序列上更为相似,可能构成了主要的DNA结合域。类群Ⅱ包含1个WRKY结构域和1个C2H2型锌指基序,并可进一步细分为5个亚群(Ⅱa~Ⅱe)。类群Ⅲ则包含1个WRKY结构域和1个以HXC结尾的C2HC型锌指基序(图1b)。3个类群WRKY转录因子的具体功能因植物种类和生长环境的不同而有所差异[14]。
2 WRKY转录因子在植物中的功能
植物在整个发育过程中会不可避免地受到各种生物和非生物胁迫[24]。研究表明,WRKY转录因子在植物应激反应及发育过程中起到关键作用。它们通过调控相关基因的表达增强植物抗逆性,为植物生长发育提供保障,尤其是在激素信号传导方面,如脱落酸(abscisic acid,ABA)、水杨酸(salicylic acid,SA)、茉莉酸(jasmonic acid,JA)等[21-25],WRKY转录因子通过调控下游基因的表达来促进或抑制相关蛋白的合成[5]。在生物胁迫和非生物胁迫过程中,WRKY转录因子可作为正调节剂或负调节剂,通过调节相关基因表达来影响植物对胁迫的耐受性。例如,在拟南芥(Arabidopsis thaliana)中,WRKY57和WRKY63基因过表达会显著提高植株抗旱性,与野生型相比,过表达植株体内的ABA含量增加,同时抵抗胁迫的相关基因表达也上调[26];大蒜(Allium sativum)中的9个AsWRKYs对盐胁迫、赤霉素胁迫和干旱胁迫均有响应[27];在药用稻(Oryza officinalis)中,OsWRKY3、OsWRKY13等转录因子可以被病原菌诱导表达,并通过调节相关基因表达参与植物的防御反应[28]。随着研究的深入,不断有新的WRKY转录因子在植物中被鉴定和分析,其功能多样性也逐渐得到阐明[29]。WRKY转录因子在不同植物中的功能预测列于表1。
表1 不同植物中WRKY转录因子的功能预测Tab.1 Functional prediction of WRKY transcription factors in different plants |
| 物种 | 鉴定数 | 功能预测 | 相应WRKY转录因子 |
|---|---|---|---|
| 大蒜(Allium sativum)[27] | 78 | 高温和高盐胁迫响应 | As29182、As40049、As20748、As31693、As36680、As38227、As39900、As46742、As91121 |
| 高粱(Sorghum bicolor)[26] | 94 | 抗干旱和保持绿色 | SbWRKY45、SbWRKY79、SbWRKY74、 SbWRKY72、SbWRKY75 |
| 拟南芥(Arabidopsis thaliana)[26] | 74 | 抗干旱和抗渗透胁迫 | AtWRKY63、AtWRKY57、AtWRKY54 |
| 茄子(Solanum melongena)[19] | 50 | 参与非生物和生物胁迫、病原菌抗性 | Smel_Unigene_26604_orf |
| 水茄(Solanum torvum)[19] | 62 | 参与非生物和生物胁迫、病原菌抗性 | Stor_Unigene_36980_orf |
| 甘蓝(Brassica oleracea)[30] | 148 | 抗逆、参与植物生长和形态发育 | BolWRKY61、BolWRKY67、BolWRKY83、 BolWRKY84、BolWRKY119、BolWRKY122、 BolWRKY117 |
| 小麦(Triticum aestivum)[31] | 172 | 热胁迫和干旱胁迫响应 | TaWRKY014、TaWRKY090、TaWRKY008、 TaWRKY122 |
| 闽楠(Phoebe bournei)[32] | 60 | 抗干旱和涝渍 | PbWRKY36 |
| 玉米(Zea mays)[33] | 106 | 高温胁迫和干旱胁迫响应、ABA信号 | ZmWRKY106 |
| 甜橙(Citrus sinensis)[34] | 47 | 抗指状青霉感染 | CsWRKY2、CsWRKY14 |
| 陆地棉(Gossypium hirsutum)[35] | 15 | 参与生长发育和叶片衰老 | GhWRKY17 |
| 药用野生稻(Oryza officinalis Wall)[28] | 89 | 参与生长发育、抗细菌和真菌 | OoWRKY13、OoWRKY71、OoWRKY111、 OoWRKY26、OoWRKY53、OoWRKY30、 OoWRKY3、OoWRKY13 |
| 甘蔗(Saccharum spontaneum)[36] | 154 | 参与糖代谢和光合作用 | SsWRKY71、SsWRKY145、SsWRKY68、 SsWRKY122、SsWRKY125、SsWRKY21、 SsWRKY22、SsWRKY39、SsWRKY56 |
| 马褂木(Liriodendron chinense)[37] | 44 | 低温、高温和干旱胁迫响应 | LchiWRKY5、LchiWRKY23、LchiWRKY14、LchiWRKY27、LchiWRKY36 |
| 亚麻(Linum usitatissimum)[38] | 105 | 参与生长发育和逆境应答 | Lus10001265、Lus10012215、Lus10042243、 Lus10026409、Lus10002309、Lus10024074、 Lus10026082 |
| 芹菜(Apium graveolens)[39] | 69 | 抗低温和干旱胁迫 | AgWRKY50、AgWRKY39、AgWRKY47、 AgWRKY71、AgWRKY8、AgWRKY2、AgWRKY38 |
| 文冠果(Xanthoceras sorbifolium)[40] | 65 | 响应盐胁迫和干旱胁迫以及激素处理 | XsWRKY20、XsWRKY34 |
| 莲花(Nelumbo nucifera)[41] | 65 | 参与生物和非生物胁迫、提高苯基异喹啉生物碱合成 | NnWRKY40a、NnWRKY40b |
| 甘薯(Ipomoea batatas)[42] | 79 | 增强盐胁迫耐受性 | IbWRKY47、IbWRKY33 |
| 菜豆(Phaseolus vulgaris)[43] | 88 | 干旱胁迫响应 | PvWRKY1、PvWRKY2、PvWRKY3、PvWRKY5、 PvWRKY21、PvWRKY24、PvWRKY28、 PvWRKY38、PvWRKY40、PvWRKY52、 PvWRKY53、PvWRKY43、PvWRKY58 |
| 半夏(Pinellia ternata)[44] | - | 高温胁迫响应 | PtWRKY2 |
从表1可以看出,WRKY转录因子在植物中扮演着多样化的角色,它们不仅通过调控激素信号传导影响植物的生长发育和逆境响应,还参与其他生理过程。WRKY转录因子调控的植物次生代谢物包括萜类、酚类和生物碱等多种化合物,这些代谢物在植物适应环境和抵御胁迫方面发挥着极其重要的作用。
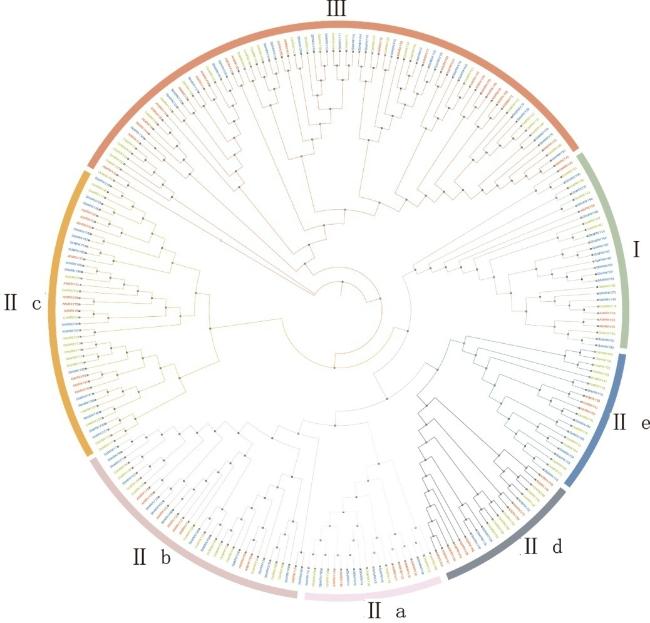
为了探究不同物种之间WRKY家族基因的系统发育和进化关系,采用MAFFT软件对拟南芥、高粱和水稻的WRKY编码氨基酸序列进行比对,利用MEGA 11.0 软件构建系统发育树(图2)。结果表明:拟南芥中参与干旱和渗透胁迫调控的AtWRKY57属于Ⅲ组,而AtWRKY63、AtWRKY54属于Ⅱd亚组,参与脱落酸信号传导和非生物胁迫的AtWRKY8和AtWRKY50属于Ⅲ组[26];水稻中与病原菌抗性相关的OsWRKY3位于Ⅱb亚组,而激活类黄酮生物合成的OsWRKY13则属于Ⅱe亚组[28];高粱中抗干旱的SbWRKY45位于Ⅰ组,SbWRKY74归属于Ⅱc亚组,SbWRKY75位于Ⅱe亚组,而SbWRKY79、SbWRKY72均属于Ⅲ组,可以调节脱落酸的SbWRKY8位于Ⅱc亚组,SbWRKY45位于Ⅰ组[26]。可以发现,调节植物的WRKY转录因子在WRKY基因家族的3个类群中均有分布。这些结果表明,与代谢和抗逆性相关的WRKY转录因子并未严格分布于WRKY基因家族的特定类群,而是广泛存在于不同类群中,反映了WRKY基因在不同物种间的进化多样性与功能复杂性。
3 WRKY转录因子在植物次生代谢过程中的调控作用
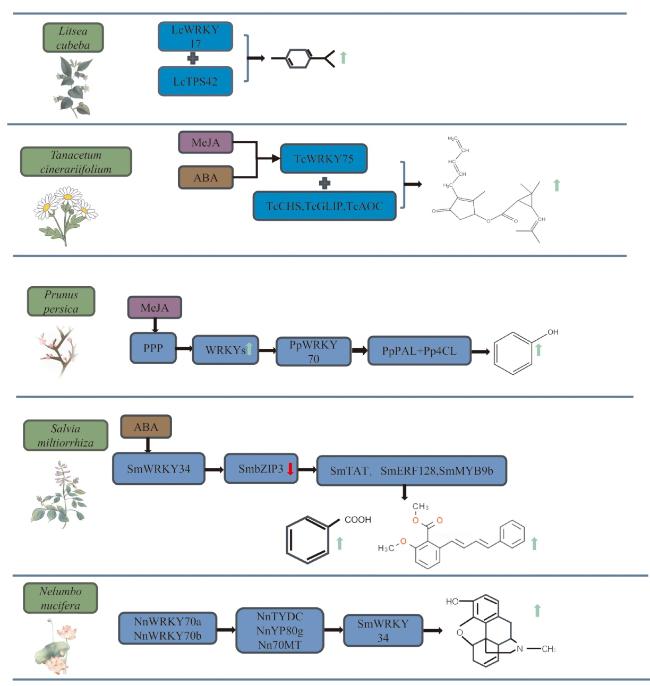
植物在长期进化过程中形成了一系列适应性产物,即次生代谢物,包括萜类、酚类和生物碱等,这些化合物在植物应对非生物胁迫(例如干旱胁迫、高温胁迫、低温胁迫、盐胁迫)和生物胁迫(例如病原菌感染和植食性昆虫侵害)中扮演着至关重要的角色。萜类化合物不仅参与胁迫信号的传递,还会吸引授粉者、捕食者和寄生性天敌[45]。酚类化合物(如多酚和黄酮类)具有强抗氧化能力,可有效清除活性氧,维持细胞内氧化还原平衡。此外,某些酚类代谢物还能抑制病原菌的酶活性并阻止其侵染[46]。生物碱类次生代谢物具有广泛的抗菌、抗病毒和抑制昆虫活性等功能。这些次生代谢物的合成和积累受到精细调控,其中转录因子扮演着关键角色。WRKY转录因子作为植物特异性转录调控网络的核心组成部分,在调控次生代谢途径中具有重要作用。不同植物的WRKY家族成员在次生代谢途径中的功能存在显著差异,这种调控多样性反映了WRKY家族成员的功能特异性及其在不同环境条件下的适应性(表2,图3)。
表2 WRKY转录因子参与的次生代谢物生物合成调控Tab.2 Biosynthesis regulation of secondary metabolites involved by WRKY transcription factors |
| 物种 | WRKY转录因子 | 次生代谢物 | 转录因子对次生代 谢物合成的作用 | WRKY转录因子 的作用位点 |
|---|---|---|---|---|
| 砂仁(Amomum villosum)[47] | AvWRKY61、AvWRKY28、AvWRKY40 | 萜烯 | 促进 | AvNeoD |
| 山鸡椒(Litsea cubeba)[48] | LcWRKY17 | 单萜类 | 促进 | LcTPS42 |
| 丹参(Salvia miltiorrhiza)[49] | SmWRKY1、SmWRKY7、SmWRKY19、SmWRKY29、SmWRKY45、SmWRKY52、SmWRKY56、SmWRKY58、SmWRKY68 | 丹参酮、酚酸 | 促进 | SmDXS2、SmGGPPS、 SmCPS、SmKSL、 SmCYP76H1 |
| 番茄(Solanum lycopersicum)[50] | SlWRKY4、SlWRKY31、SlWRKY37 | 总酚、类黄酮 | 促进 | β-葡聚糖酶A、防御素、几丁质酶 |
| 葡萄(Vitis vinifera)[51] | VvWRKY8 | 白藜芦醇 | 抑制 | VvSTS15、VvSTS21 |
| 小蛇根草(Ophiorrhiza pumila)[52] | OpWRKY2 | 喜树碱 | 促进 | OpTDC |
| 苹果(Malus domestica)[53] | MdWRKY75、MdWRKY1 | 花青素 | 促进 | MdLNC499、MdERF109 |
| 马铃薯(Solanum tuberosum)[54] | StWRKY70 | 花青素 | 促进 | StDFR、StAN1 |
| 红豆杉(Taxus chinensis)[55] | TcWRKY33 | 紫杉醇 | 促进 | TcDBAT、TcTASY |
| 青蒿(Artemisia annua)[56] | AaWRKY9 | 青蒿素 | 促进 | AaDBR2、AaGSW1 |
| 烟草(Nicotiana tabacum)[57] | NtWRKY3 | 东莨菪碱、小檗碱 | 促进 | NtLOX3、NtACS1、NtACO1 |
| 桔梗(Platycodon gran- diflorus)[58] | PgWRKY2、PgWRKY9、PgWRKY10、PgWRKY24 | 三萜皂苷 | 促进 | PgHMGS、PgHMGR、PgMK、PgMVD |
| 桃树(Prunus persica)[59] | PpWRKY70 | 绿原酸 | 促进 | PpPAL、Pp4CL |
| 啤酒花(Humulus lupulus)[60] | HlWRKY1 | 异戊烯基黄酮 | 促进 | HlWDR1 |
| 灯盏花(Erigeron breviscapus)[61] | EbWRKY11、EbWRKY36、 EbWRKY44 | 黄酮类化合物 | 促进 | EbF6H |
| 百香果(Passiflora edulis)[62] | PeWRKY48 | 黄酮类化合物 | 促进 | PeF3H、PeCHI |
| 除虫菊(Tanacetum cinerariifolium)[63] | TcWRKY75 | 除虫菊酯 | 促进 | TcCHS、TcAOC、TcGLIP |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3.1 WRKY对植物萜类化合物合成的影响
萜类也被称为类异戊二烯,是由异戊二烯单元衍生而成的一类极为丰富的天然产物。根据异戊二烯单元的数量,萜类可分为单萜、倍半萜、二萜、三萜及多萜[64-66]。迄今为止,已鉴定的萜类化合物超过8万种,它们在医疗、环保和食品等领域扮演着多重角色,包括药用、植保防控、食品保鲜等。例如,萜类化合物可作为化感物质调节植物生长,增强植物的抗病能力,或通过抑制病原微生物的生长提高植物免疫力。萜类用于食品中的致病菌防治,可延长食品保鲜期[67-68]。萜类化合物的生物合成主要涉及2条代谢途径,即甲羟戊酸(mevalonate,MVA)途径和甲基赤藓糖醇磷酸(methylerythritol phosphate,MEP) 途径,分别在细胞质和过氧化物酶体及质体中进行[69-71]。
近年来,研究证明了WRKY转录因子在植物萜类合成中的重要作用。例如,山苍子中的64个WRKY基因分布于12条染色体上,其中LcWRKY17特异性调控单萜类化合物的合成。通过双荧光素酶和酵母单杂交分析,证实LcWRKY17通过结合LcTPS42启动子的W-box元件激活其转录,从而促进单萜合成[46]。除虫菊以其产生的具有杀虫作用的单萜衍生物除虫菊酯著称[72-73],其腺体定位、响应茉莉酸甲酯(methyl jasmonate,MeJA)处理的TcWRKY75转录因子能够通过结合除虫菊酯合成基因TcCHS、TcGLIP以及 TcAOC启动子的W-box元件,促进除虫菊酯的合成与积累[63]。黄花蒿全基因组分析鉴定出122个WRKY基因,其中AaWRKY40启动子区包含多个非生物胁迫顺式调控元件,AaWRKY40与AaWRKY1在茉莉酸甲酯处理的黄花蒿细胞培养物中的表达分析结果表明,AaWRKY40可能参与青蒿素的合成[74]。人参中的PgWRKY4X能够与角鲨烯环氧化酶启动子结合,提高人参皂苷生物合成相关基因的表达,促进人参皂苷积累[75]。西洋参中的PqWRKY1在拟南芥中异源过表达能够激活三萜生物合成途径[76];在桔梗中,PgWRKY2、PgWRKY9、PgWRKY10、PgWRKY24这4个转录因子与三萜生物合成途径基因正相关,这些基因分布在系统发育类群Ⅰ和Ⅱ中,这与西洋参中的研究结果一致[58]。在香椿(Toona sinensis)中,共表达网络分析揭示了78个TsWRKY基因中的多个成员与10个萜烯合成基因的表达高度相关,尤其是TsFPPS、TsIDI、TsMTPS等,TsWRKY9、TsWRKY24、TsWRKY35可能协同调节萜烯合成[77]。
3.2 WRKY对植物酚类化合物合成的影响
研究表明,WRKY转录因子在植物酚类次生代谢物的合成和调节中起重要作用,这些转录因子通过激活与抗逆相关的代谢途径来影响植物中的酚类物质含量。例如,在小麦中8个TaWRKY基因在盐胁迫下表现出显著诱导作用,尤其是TaWRKY6的表达量在Sids 14和Sakha 93中急剧增加,同时小麦中的总糖、脯氨酸和酚类物质含量显著升高[84]。MeJA通过调控苯丙烷途径,启动桃树根霉防御的分子机制,特别是通过启动PpWRKY70表达,激活PpPAL和Pp4CL启动子,提高这些酶的活性及总酚类、总黄酮的含量,从而增强植株抗病性,降低病害发生率[59]。在高温胁迫下,4个莴苣品种中鉴定到大量与类黄酮生物合成相关的代谢物和差异表达基因,此外,鉴定的31个转录因子中有25个上调,其中WRKY转录因子在类黄酮合成中起到重要作用[52]。蒲公英(Taraxacum antungense)中含有的木犀草素是蒲公英的主要次生代谢物,它是一种低分子量的多酚类化合物,TaWRKY44基因表达上调时,蒲公英中木犀草素的含量增加[85]。在黄芩(Scutellaria baicalensis)中,WRKY转录因子在干旱胁迫下的表达调控呈现复杂模式,在中度干旱胁迫下,与黄芩苷合成密切相关的某些WRKY转录因子(如SbWRKY8和SbWRKY16)表达显著上调,促进黄芩苷合成;而在严重干旱胁迫下,WRKY家族中部分成员的表达显著降低,黄芩苷的合成也受到抑制[86]。在丹参中,SmWRKY34通过直接调控关键基因,如SmRAS和SmGGPPS,以及通过ABA响应性的bZIP转录因子SmbZIP3,调节丹参酮和酚酸的合成[87]。
3.3 WRKY对植物生物碱合成的影响
生物碱的合成和调控同样受到WRKY转录因子的影响。例如,在黄连(Coptis chinensis)中,鉴定出 41个与原小檗碱生物合成相关的 WRKY 转录因子,基于基因表达模式、代谢途径、系统发育和双荧光素酶分析,发现CcWRKY7、CcWRKY29 和CcWRKY32能够调控原小檗碱的生物合成[94]。日本海棠(Chaenomeles japonica)中的CjWRKY1被认为是苯甲基异喹啉生物碱(benzylisoquinoline alkaloid,BIA)合成途径中的关键转录因子,其在花菱草(Eschscholzia californica)中的过表达可上调多种BIA合成酶基因的表达,并提高BIA的积累[95]。在花菱草中,鉴定出50个EcWRKY基因,部分EcWRKY基因能够响应MeJA,且这些基因的表达模式与BIA生物合成酶基因的表达模式相似,可能在BIA合成中发挥作用[96]。在荷花(Nelumbo nucifera)中,NnWRKY70b通过激活包括NnTYDC、NnYP80g、Nn7OMT在内的BIA合成基因启动子,促进BIA积累;而NnWRKY70a仅能够激活NnTYDC的启动子。瞬时过表达实验表明,NnWRKY70a和NnWRKY70b显著提高了BIA在莲花花瓣中的积累,其中NnWRKY70b的调控作用更强[97]。在马铃薯(Solanum tuberosum)中,StWRKY8对BIA的合成起正调控作用,特别是在抗病基因型中,StWRKY8与BIA生物合成基因的启动子结合,促进抗菌化合物积累,并增强细胞壁的防御作用[98]。在小蛇根草(Ophiorrhiza pumila)中鉴定出46个OpWRKY基因,这些基因不均匀分布于11条染色体上。过表达OpWRKY6会显著减少喜树碱的积累,而OpWRKY6敲除则可以显著增加喜树碱含量。OpWRKY6通过直接下调OpGES、Op10HGO、Op7DLH和OpTDC基因的表达,负向调控喜树碱的生物合成[99]。在短小蛇根草(Ophiorrhiza pumila)中,OpWRKY1主要在茎中表达,过表达OpWRKY1可抑制喜树碱合成相关基因如OpCPR的表达,显著减少喜树碱积累[100]。在长春花(Catharanthus roseus)中,CrWRKY1受到茉莉酸、赤霉酸和乙烯的诱导,优先在根中表达。CrWRKY1通过与色胺脱羧酶(tryptophan decarboxylase,TDC)启动子中的W-box结合,促进TDC表达并增加色胺浓度,从而提高吲哚生物碱类化合物的积累[101]。
4 结论
尽管WRKY转录因子在调控植物次生代谢途径中的作用已被广泛研究,但现有研究多集中于特定的几种模式植物和有限的次生代谢物,缺乏广泛的物种覆盖和系统深入的功能解析,这限制了对WRKY转录因子在广泛植物物种中作用的全面理解。此外,现有研究往往侧重于WRKY家族中的某一种或少数成员,忽视了其他成员的可能作用及成员间的相互作用,详细的分子机制和调控网络尚未完全建立,导致对WRKY转录因子家族功能的认识还不够全面。同时,WRKY转录因子家族与其他转录因子如MYB、bHLH等的相互作用及其在复杂生理和代谢过程中的角色也尚未充分阐明。
未来研究可考虑利用高通量组学技术、生物信息学方法以及CRISPR等基因编辑工具,全面系统解析WRKY家族成员的功能及其在多物种中的作用,包括它们如何通过与其他转录因子及信号分子的交互作用来调控植物次生代谢物的合成。通过现代分子生物学技术,揭示WRKY转录因子在非模式植物中的功能,探索其调控网络和作用机制,揭示其在植物次生代谢物合成中的具体作用。此外,应注重WRKY转录因子在特定组织或发育阶段的表达调控研究,以精确平衡植物生长与次生代谢物积累之间的关系。利用上述深入研究,不仅能够更全面地理解WRKY转录因子在植物次生代谢中的调控机制,还能开发新策略,利用分子育种或基因工程手段提高作物的次生代谢物合成和抗逆性,显著优化植物的农艺性状和经济价值,为食品安全和可持续农业生产提供坚实的科学基础。