

1 材料与方法
1.1 实验材料
1.2 动物饲养及常规设计
表1 基础日粮组成及营养水平(风干基础)Tab.1 Composition and nutrient levels of the basal diet(air-dry basis) |
| 成分 | 组 别 | ||||
|---|---|---|---|---|---|
| Con | G3 | G6 | G9 | G12 | |
| 玉米青贮 | 67.60 | 64.60 | 62.20 | 58.40 | 57.30 |
| 小麦秸秆 | 10.00 | 10.08 | 9.80 | 9.60 | 9.47 |
| 玉米 | 8.18 | 9.18 | 9.58 | 9.98 | 10.31 |
| 豆粕 | 6.32 | 5.34 | 4.72 | 4.42 | 5.16 |
| 菜粕 | 2.42 | 2.92 | 2.82 | 2.22 | 0.88 |
| 银杏叶粉 | 0.00 | 3.00 | 6.00 | 9.00 | 12.00 |
| 小麦麸 | 2.48 | 1.88 | 1.88 | 3.38 | 1.88 |
| 食盐 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| 磷酸氢钙 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| 预混料 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| 代谢能 | 10.08 | 10.07 | 10.06 | 10.06 | 10.06 |
| 粗蛋白 | 11.16 | 11.13 | 11.11 | 11.18 | 11.13 |
| 粗脂肪 | 3.46 | 3.33 | 3.21 | 3.09 | 2.97 |
| 灰分 | 4.46 | 4.65 | 4.84 | 5.04 | 5.21 |
| 中性洗涤纤维 | 40.13 | 40.33 | 40.59 | 40.82 | 41.02 |
| 酸性洗涤纤维 | 24.07 | 24.17 | 24.20 | 24.05 | 24.17 |
| 钙 | 0.54 | 0.53 | 0.52 | 0.51 | 0.50 |
| 磷 | 0.51 | 0.50 | 0.49 | 0.49 | 0.46 |
注:每千克预混料提供维生素A 100 kIU,维生素D 80 kIU,维生素E 500 IU,Fe 760 mg,Zn 500 mg,Cu 200 mg,Mn 280 mg。代谢能单位为MJ/kg,其他单位为%。 |
1.3 样品采集及指标测定
1.3.1 生长性能
1.3.2 表观消化率
1.3.3 血液采集
1.3.4 瘤胃液采集
1.3.5 血清生化指标和血清抗氧化指标的测定
1.3.6 瘤胃液pH值、氨态氮及挥发性脂肪酸测定
1.3.7 瘤胃液总菌DNA的提取及16S rRNA测序
1.3.8 瘤胃液代谢组学分析
1.4 数据统计与分析
2 结果与分析
2.1 银杏叶粉对安格斯牛生长性能的影响
表2 银杏叶粉对安格斯牛生长性能的影响Tab.2 Effect of Ginkgo biloba powder on the growth performance of Angus cattle |
| 指标 | 组 别 | 标准 误差 | P值 | ||||
|---|---|---|---|---|---|---|---|
| Con | G3 | G6 | G9 | G12 | |||
| 初重/kg | 459.00 | 449.25 | 454.50 | 452.20 | 447.25 | 4.39 | 0.96 |
| 末重/kg | 539.80 | 539.50 | 546.60 | 548.80 | 538.00 | 4.55 | 0.96 |
| 平均日增重/(kg·d-1) | 1.14 | 1.22 | 1.25 | 1.27 | 1.22 | 0.26 | 0.09 |
| 干物质采食量/(kg·d-1) | 10.51b | 11.52a | 11.68a | 12.23a | 11.60a | 0.14 | 0.02 |
| 料重比 | 9.34 | 9.57 | 9.27 | 9.61 | 9.34 | 0.19 | 0.92 |
注:同行数据肩标不同小写字母表示差异显著(P<0.05)。 |
2.2 银杏叶粉对安格斯牛表观消化率的影响
表3 银杏叶粉对安格斯牛表观消化率的影响Tab.3 Effect of Ginkgo biloba powder on the apparent digestibility of Angus cattle |
| 指标 | 组 别 | 标准 误差 | P值 | ||||
|---|---|---|---|---|---|---|---|
| Con | G3 | G6 | G9 | G12 | |||
| 干物质消化率/% | 73.56 | 75.02 | 75.92 | 76.42 | 75.36 | 0.72 | 0.80 |
| 粗蛋白消化率/% | 70.88b | 73.34ab | 78.97a | 78.28a | 75.11ab | 0.87 | 0.03 |
| 粗脂肪消化率/% | 58.30 | 60.38 | 63.13 | 64.96 | 65.09 | 1.01 | 0.13 |
| 中性洗涤纤维消化率/% | 56.06c | 58.84bc | 63.16ab | 66.12a | 60.12abc | 0.93 | 0.01 |
| 酸性洗涤纤维消化率/% | 36.27b | 42.83ab | 45.44a | 48.60a | 48.21a | 1.20 | 0.04 |
注:同行数据肩标不同小写字母表示差异显著(P<0.05)。 |
2.3 银杏叶粉对安格斯牛血清生化指标和抗氧化指标的影响
表4 银杏叶粉对安格斯牛血清生化指标的影响Tab.4 Effect of Ginkgo biloba powder on serum biochemical indexes of Angus cattle |
| 指标 | 组 别 | 标准 误差 | P值 | ||||
|---|---|---|---|---|---|---|---|
| Con | G3 | G6 | G9 | G12 | |||
| 总蛋白/ (g·L-1) | 74.80 | 71.53 | 75.35 | 77.10 | 72.25 | 1.38 | 0.74 |
| 白蛋白/ (g·L-1) | 28.48 | 29.00 | 29.15 | 30.33 | 28.35 | 0.42 | 0.64 |
| 谷丙转氨酶/(U·L-1) | 29.50 | 32.15 | 32.48 | 36.88 | 33.85 | 1.06 | 0.28 |
| 谷草转氨酶/(U·L-1) | 84.48 | 81.88 | 76.50 | 81.43 | 81.50 | 2.16 | 0.87 |
| 总胆固醇/(mmol·L-1) | 4.19 | 3.67 | 3.49 | 3.48 | 3.34 | 0.51 | 0.13 |
| 葡萄糖/(mmol·L-1) | 3.54 | 4.03 | 3.83 | 3.78 | 3.55 | 0.09 | 0.43 |
| 甘油三酯/(mmol·L-1) | 0.28a | 0.25ab | 0.24ab | 0.20b | 0.21b | 0.01 | 0.04 |
| 尿素氮/(mmol·L-1) | 6.28 | 5.94 | 6.85 | 6.13 | 6.50 | 0.29 | 0.91 |
| Ca2+/(mmol·L-1) | 2.52ab | 2.52ab | 2.57ab | 2.68a | 2.39b | 0.03 | 0.04 |
注:同行数据肩标不同小写字母表示差异显著(P<0.05)。 |
表5 银杏叶粉对安格斯牛血清抗氧化指标的影响Tab.5 Effect of Ginkgo biloba powder on serum antioxidant indexes of Angus cattle |
| 指标 | 组 别 | 标准 误差 | P值 | ||||
|---|---|---|---|---|---|---|---|
| Con | G3 | G6 | G9 | G12 | |||
| 总抗氧化能力/ (μmol·mL-1) | 7.70b | 8.62ab | 8.96ab | 9.50a | 7.94b | 0.22 | 0.03 |
| 谷胱甘肽过氧化物酶/ (U·mL-1) | 97.44 | 114.73 | 117.92 | 105.03 | 96.32 | 3.07 | 0.06 |
| 超氧化物歧化酶/ (U·mL-1) | 125.00 | 137.25 | 138.25 | 141.50 | 133.25 | 3.18 | 0.57 |
| 丙二醛/ (μmol·L-1) | 6.56a | 6.31a | 5.09b | 5.38b | 5.65b | 0.15 | <0.01 |
注:同行数据肩标不同小写字母表示差异显著(P<0.05)。 |
2.4 银杏叶粉对安格斯牛瘤胃发酵参数的影响
表6 银杏叶粉对安格斯牛瘤胃发酵参数的影响Tab.6 Effect of Ginkgo biloba powder on rumen fermentation parameters of Angus cattle |
| 参数 | 组 别 | 标准 误差 | P值 | ||||
|---|---|---|---|---|---|---|---|
| Con | G3 | G6 | G9 | G12 | |||
| pH值 | 6.88 | 6.93 | 7.14 | 7.05 | 6.93 | 0.38 | 0.44 |
| 氨态氮/(mg·dL-1) | 11.20 | 10.89 | 10.81 | 10.04 | 10.69 | 0.16 | 0.29 |
| 总挥发性脂肪酸/(mmol·L-1) | 70.20 | 67.36 | 66.19 | 67.48 | 68.39 | 0.53 | 0.22 |
| 乙酸/% | 67.83a | 65.61c | 64.44c | 65.86bc | 67.35ab | 0.34 | <0.01 |
| 丙酸/% | 17.99c | 18.56c | 21.71a | 20.08b | 18.78c | 0.31 | <0.01 |
| 异丁酸/% | 1.16 | 1.11 | 0.98 | 1.08 | 1.02 | 0.05 | 0.53 |
| 丁酸/% | 8.10b | 10.12a | 8.29b | 8.34b | 8.61b | 0.21 | <0.01 |
| 异戊酸/% | 3.20 | 2.98 | 2.78 | 2.82 | 2.67 | 0.07 | 0.15 |
| 戊酸/% | 1.71 | 1.64 | 1.80 | 1.81 | 1.56 | 0.03 | 0.09 |
注:同行数据肩标不同小写字母表示差异显著(P<0.05)。 |
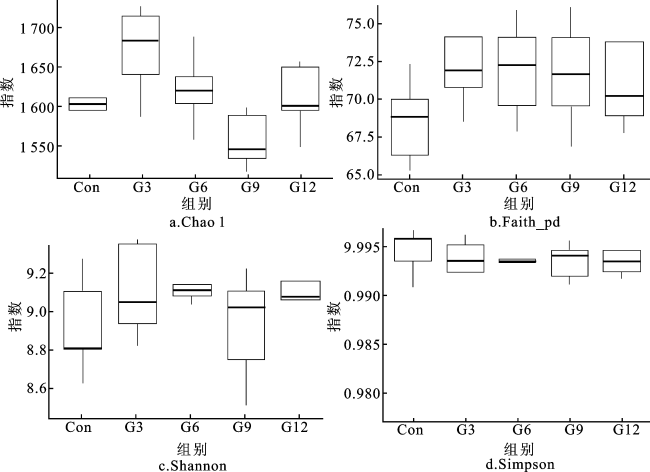
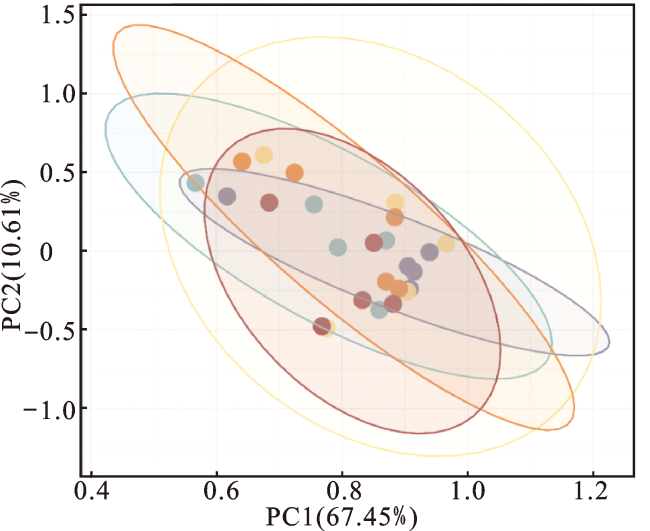
2.5 银杏叶粉对安格斯牛瘤胃菌群丰富度和多样性的影响
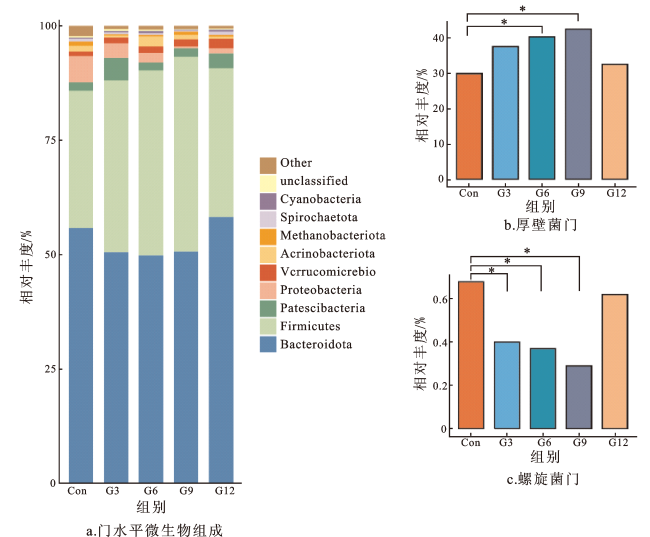
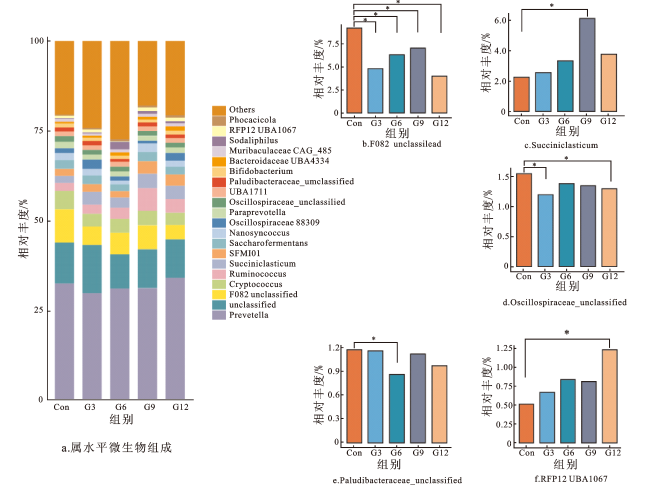
2.6 银杏叶粉对安格斯牛瘤胃微生物门、属水平相对丰度的影响
2.7 瘤胃微生物功能预测
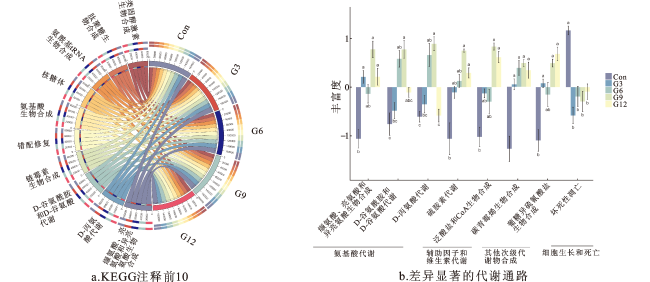
2.8 银杏叶粉对安格斯牛瘤胃代谢物的影响
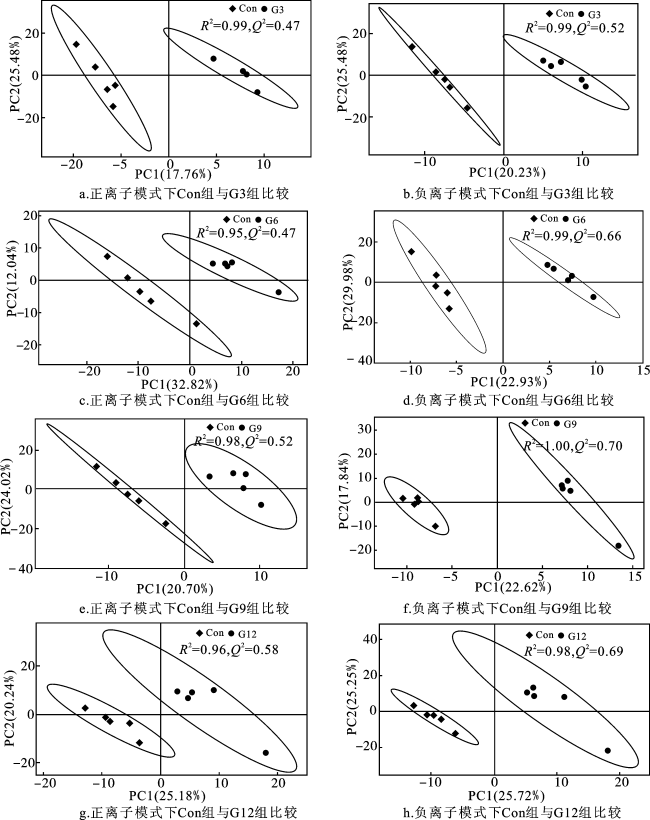
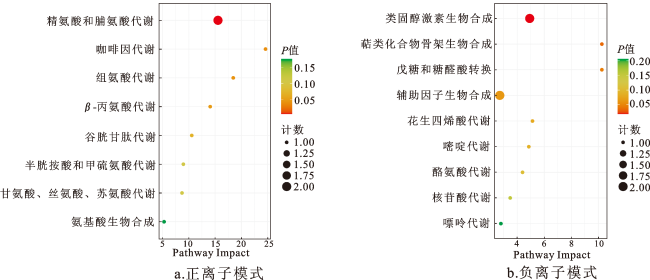
表7 正离子模式下组间差异代谢物对比分析(前10)Tab.7 Comparative analysis of differential metabolites between groups in positive ion mode(top 10) |
| 代谢物 | 生物角色 | 组 别 | VIP | P值 | ||||
|---|---|---|---|---|---|---|---|---|
| Con | G3 | G6 | G9 | G12 | ||||
| 胆固醇酯 | 甾醇脂类 | 1.58b | 2.57a | 3.07a | 3.20a | 1.26b | 2.82 | <0.01 |
| N-乙酰缬氨酸 | 有机酸 | 2.23c | 3.28b | 3.56b | 4.36a | 3.33b | 2.81 | <0.01 |
| (9Z,11E,13S,15Z)-13-羟 基十八烷-9,11,15-三烯酸 | 脂肪酸 | 14.64a | 8.82bc | 7.58c | 7.01c | 12.93ab | 2.39 | <0.01 |
| 1-(3,4-二甲氧基苯基)-乙酮肟 | 其他 | 0.24b | 0.36a | 0.37a | 0.39a | 0.32ab | 2.37 | 0.02 |
| 油酸乙酯 | 脂肪酸 | 0.21ab | 0.16ab | 0.11c | 0.09c | 0.24a | 2.37 | 0.03 |
| 异丙基β-D-吡喃葡萄糖苷 | 碳水化合物 | 0.07b | 0.93a | 0.97a | 1.28a | 0.80ab | 2.32 | 0.04 |
| 1,7-双(4-羟基苯基)-3-庚酮 | 多酚类物质 | 0.24c | 0.40b | 0.61a | 0.68a | 0.33bc | 2.22 | <0.01 |
| 四醇-12R-HETE | 其他 | 3.61c | 12.68c | 21.74bc | 39.62ab | 51.12a | 2.20 | <0.01 |
| N-丁基-2-甲基-5-(哌 啶基磺酰基)-3-呋喃酰胺 | 其他 | 1.19b | 1.21b | 1.48a | 1.48a | 1.27ab | 2.15 | 0.04 |
| 4-氧代-4-[(1-苯乙 基)氨基]丁-2-烯酸 | 有机氮化合物 | 4.09c | 4.03c | 8.28ab | 10.02a | 5.96bc | 2.14 | 0.04 |
注:VIP为变量投影重要性。同行数据肩标不同小写字母表示差异显著(P<0.05)。 |
表8 负离子模式下组间差异代谢物对比分析(前10)Tab.8 Comparative analysis of differential metabolites between groups in negative ion mode(top 10) |
| 代谢物 | 生物角色 | 组 别 | VIP | P值 | ||||
|---|---|---|---|---|---|---|---|---|
| Con | G3 | G6 | G9 | G12 | ||||
| 8-异-15-酮前列腺素E2 | 激素和递质类 | 0.10d | 0.45c | 0.93a | 0.65b | 0.15d | 3.87 | <0.01 |
| 6-十五酰基水杨酸 | 苯及其取代衍生物 | 0.03c | 0.53bc | 0.94ab | 1.53a | 1.56a | 3.09 | <0.01 |
| 20-羟基二十碳四烯酸 | 脂肪酸 | 0.10c | 1.99bc | 3.10b | 6.18a | 6.11a | 3.00 | <0.01 |
| 玉米赤霉醇 | 大环内酯类和类似物 | 3.05c | 11.02bc | 21.68ab | 35.33a | 37.97a | 2.66 | <0.01 |
| 7-酮胆酸 | 类固醇 | 0.02c | 0.09c | 0.17b | 0.30ab | 0.40a | 2.54 | <0.01 |
| 戊二酸 | 羧酸及其衍生物 | 13.03b | 24.64a | 25.65a | 25.05a | 17.98ab | 2.54 | 0.01 |
| 乳清酸 | 二嗪 | 0.32a | 0.30ab | 0.18c | 0.19bc | 0.30ab | 2.46 | 0.02 |
| L-阿拉伯糖醇 | 有机氧化合物 | 0.39a | 0.29ab | 0.08b | 0.10b | 0.19ab | 2.32 | <0.01 |
| 乌头酸 | 萜类化合物 | 74.41b | 73.49b | 88.91a | 90.27a | 67.62b | 2.21 | 0.04 |
| 17(S)-HP二氢乙酸 | 脂肪酸 | 0.03c | 0.06c | 0.13b | 0.23ab | 0.31a | 2.17 | <0.01 |
注:VIP为变量投影重要性。同行数据肩标不同小写字母表示差异显著(P<0.05)。 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}