

葡萄(Vitis vinifera)原产于亚洲西部,是世界上最古老的果树之一,不仅营养价值高,在中药中也应用广泛,是享誉世界的营养、药用商品[1]。中国是世界葡萄属植物的重要起源地和分布中心之一,栽培面积广,主栽培区在北纬25°~45°范围内,但北方的干旱严寒气候制约了鲜食葡萄及酿酒葡萄的大规模生产和产量提升[2-3]。目前,杂交育种是解决葡萄抗寒和抗旱的主要措施,但杂交葡萄具有优良性状相互连锁、育种周期长、工作量大等问题,传统育种并不能满足葡萄产业化的需求[2]。近年来,随着多个葡萄品种全基因组测序的完成,许多优良的抗逆基因资源被发掘,因此在分子调控机制上对葡萄育种方式进行改良愈发表现出优势[4⇓⇓-7]。
葡萄抗寒性和抗旱性涉及基因众多,根据其作用方式可简单归纳为调控基因和功能基因2类。调控基因主要通过调控抗性基因的表达、逆境信号感知与传导、转录调控及蛋白水平修饰等过程来提高植物的抗逆性,如WRKY转录因子、CBF转录因子、DREB转录因子、MADS-box转录因子家族等;而功能基因则与植物抗寒性或抗旱性的提高直接相关,如抗冻蛋白基因、抗氧化酶基因、渗透调节蛋白基因等[8⇓⇓⇓-12]。
MADS-box转录因子家族是真核生物中最重要的转录因子家族之一,在动植物和真菌中被广泛研究。该家族的特征是有1个保守性很强的DNA结合结构域[13]。植物中MADS-box家族分为Type-Ⅰ型和Type-Ⅱ型,其中关于Type-Ⅰ型的研究很少。Type-Ⅱ型也被称为MIKC类MADS-box,其含有4个结构域(MADS盒、Intervening-region、Keratin盒和C-terminal),根据保守结构域中是否含有Keratin盒可将其分为MIKCC和MIKC*。MIKCC是Type-Ⅱ中研究最广泛、机理较清晰的类型,其可进一步分为12个亚家族:AG-like、AGL2-like、AGL6-like、AGL12-like、AGL15-like、AGL17-like、DEF/GLO-like、FLC-like、GGM13-like、SQUA-like、STMADS11-like和TM3-like[14]。
MADS-box家族参与多种植物的生长发育与调控,不仅在花器官发育与开花时间、果实发育、植株根系发育和植株构型等过程中发挥着重要调控作用,还在响应激素与盐胁迫、干旱胁迫、低温胁迫等非生物胁迫方面发挥作用[15⇓⇓⇓⇓-20]。拟南芥(Arabidopsis thaliana)MADS-box转录因子AtAGL16通过结合CYP707A3启动子的CArG-box序列激活CYP707A3并抑制AAO3和SDD1的转录表达,从而改变叶片气孔密度、气孔运动和脱落酸(abscisic acid,ABA)含量,参与拟南芥响应干旱胁迫的应答[21]。研究发现,水稻(Oryza sativa)OsMADS23与SAPK9相互作用被磷酸化后,通过激活水稻植株中ABA合成基因和脯氨酸合成基因的表达,提高ABA与脯氨酸的积累,从而增强植株耐盐胁迫和抗旱的能力[22-23]。在水稻中,过量表达OsMADS57可维持水稻分蘖在低温胁迫下的生长,进一步研究发现OsMADS57与OsTB1共同协调其靶标OsWRKY94和D14的转录,调控水稻由器官发育转变为低温胁迫适应[24]。
AGL12-like亚组的功能仅在拟南芥、水稻和核桃中被报道过。在拟南芥中,AGL12与根系分生组织细胞增殖、花期转换和细胞周期调节有关;同时AtAGL12影响核桃体细胞胚的发芽率和根系构型[25⇓-27]。OsMADS26属于水稻AGL12亚组,在水稻和拟南芥中分别过表达该基因的转基因植物均表现出与胁迫反应相关的表型,如褪绿、细胞死亡、色素积累和根芽生长缺陷等,而在水稻中下调表达该基因的转基因植株增强了对病原菌和干旱胁迫的抗性[28]。然而,葡萄中VvAGL12启动子(proVvAGL12)的生物学功能和调控机制还有待进一步研究。
前人研究发现,基因表达受启动子区域的顺式作用元件调控[29-30]。除了TATA盒子和CAAT盒子等核心元件外,启动子还具有与激素响应、非生物胁迫响应、光响应等相关的保守特异性元件,可调节植物生长发育或响应逆境胁迫[31]。小麦(Triticum aestivum)TaIRI启动子区域含有多个低温、干旱响应元件(MYB、Myb、MYC、LTR)和茉莉酸甲酯(jasmonic acid methyl ester,MeJA)响应元件(CGTCA-motif、TGACG-motif),后续烟草(Nicotiana benthamiana)瞬时转化和拟南芥稳定转化实验证明,该启动子的活性受低温和MeJA的显著诱导[32]。腊梅(Chimonanthus praecox)CpNAC1启动子含有多个逆境胁迫相关顺式作用元件,含有该启动子的转基因拟南芥GUS活性受ABA和多种胁迫处理(4 ℃、42 ℃、NaCl、聚乙二醇)的显著诱导,进一步研究发现CpNAC1过表达拟南芥对干旱胁迫有响应[33]。
本研究从黑比诺葡萄中克隆获得VvAGL12启动子序列并分析其顺式作用元件,构建GUS融合表达载体并稳定转入拟南芥、瞬时转入烟草。通过对转基因植株进行处理,研究该启动子是否响应激素与非生物胁迫,解析其调控关系,为VvAGL12基因的生物学功能和生产应用提供参考。
1 材料与方法
1.1 实验材料
1.1.1 植物材料
实验植物材料包括葡萄测序品种黑比诺(Vitis vinifera)、拟南芥哥伦比亚型Col-0(Arabidopsis thaliana)和本氏烟草(Nicotiana benthamiana)。
1.1.2 载体与菌株
大肠杆菌(Escherichia coli,E.coli)菌株DH5α、农杆菌(Agrobacterium tumefaciens)菌株GV3101购自北京擎科生物科技有限公司。植物表达载体pCAMBIA1300+pBI101由鲁东大学农林工程研究院保存。
1.1.3 主要试剂
限制性内切酶(BamH Ⅰ、Sal Ⅰ)、DNA Marker(DL2000)、高保真酶PrimeSTAR®HS DNA Polymerase、T4连接酶购于宝生物工程(大连)有限公司;抗生素卡那霉素(Kanamycin)、利福平(Rifampicin)、潮霉素(Hygromycin)以及植物激素赤霉素(gibberellin,GA3)、脱落酸(abscisic acid,ABA)和聚乙二醇(polyethylene glycol,PEG)等常用生化试剂均购于生工生物工程(上海)股份有限公司;植物基因组DNA提取试剂盒、琼脂糖凝胶DNA回收试剂盒、质粒DNA提取试剂盒购于天根生化科技(北京)有限公司;5-溴-4-氯-3-吲哚-β-D-葡萄糖醛酸环己胺盐(X-Gluc)购于北京索莱宝科技有限公司。
1.2 实验方法
1.2.1 DNA提取与基因克隆
以黑比诺葡萄叶片为材料,使用植物基因组DNA提取试剂盒提取DNA。以葡萄基因组文库(https://www.genoscope.cns.fr/externe/GenomeBrowser/Vitis)中VvAGL12基因起始密码子上游2 kb序列为参考,设计启动子特异性引物进行PCR扩增。引物序列为proVvAGL12-F: GTCGACGTTGGATCAATTTAGTTACTTCC(下划线为BamH Ⅰ酶切位点);proVvAGL12-R: CGGATCCTGCAGACAACCTCCCACTAG(下划线为Sal Ⅰ酶切位点)。按照高保真酶说明书进行扩增,获得约1.8 kb的目的条带,胶回收产物用BamH Ⅰ和Sal Ⅰ酶切后,连接至pCAMBIA1300+pBI101载体,连接产物转化E.coli。选择单克隆送公司测序,得到正确的重组质粒P1300-proVvAGL12。
1.2.2 启动子序列分析
用数据库PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html)和PLACE(https://www.dna.affrc.go.jp/PLACE/?action=newplace)分析葡萄VvAGL12启动子(proVvAGL12)的序列,预测其响应元件。
1.2.3 拟南芥转化
将空载质粒和重组质粒P1300-proVvAGL12分别转入农杆菌GV3101,挑取单克隆PCR鉴定,将鉴定正确的菌液接种于200 mL LB液体培养基(50 mg/L Rifampicin+50 mg/L Kanamycin,文中简写为Rif+Ka),放置于28 ℃、180 r/min恒温摇床中培养至OD600 1.2~1.6。所得菌液在4 ℃、5 000 r/min条件下离心15 min,加入重悬液得到OD600 0.6~0.8的重悬菌液,用于拟南芥侵染转化。
野生型拟南芥(Col-0)种子用体积分数10%的次氯酸钠消毒10 min,12 000 r/min离心1 min,使用灭菌的超纯水冲洗5~6次,置于4 ℃春化3~5 d。将春化后的种子播种在MS培养基中,待长出4叶后将拟南芥转移至土中(m蛭石∶m营养土=1∶1),培养条件为16 h光照(22 ℃)、8 h黑暗(21 ℃)。将盛花期拟南芥剪掉果荚,花序部分倒扣于重悬菌液中浸染1.5 min,将转化后的拟南芥侧放于黑暗条件下培养24 h,之后正常培养条件培养至种子收获。将转基因拟南芥种子种植在含有抗生素(65 mg/L Hygromycin)的平板上进行筛选,抗性苗移苗至土中继续PCR验证,直到获得T3代纯合体proVvAGL12拟南芥,作为后续实验材料。
1.2.4 激素和非生物胁迫处理下VvAGL12启动子在拟南芥中的表达活性
将消毒春化后的转基因拟南芥种子播种在MS平板上,生长9 d后对拟南芥分别进行对照(CK)、激素(GA3和ABA)和非生物胁迫(PEG和低温4 ℃)处理。分别配置10 mg/L GA3,0.01 mmol/L ABA和10% PEG6000处理液,直接喷洒在拟南芥幼苗上,低温处理将幼苗放置于4 ℃,处理4 h时取拟南芥幼苗进行GUS染色分析。
1.2.5 激素和非生物胁迫处理下VvAGL12启动子在烟草表皮细胞中的表达活性
烟草种子直接撒播在土中(m蛭石∶m营养土=1∶1),4叶期时将烟草幼苗移苗至新的花盆中(每盆1棵),25 ℃、16 h光照培养约30 d,用于瞬时表达分析。
将携带空载质粒和重组质粒P1300-proVvAGL12的农杆菌GV3101置于28 ℃、180 r/min的恒温摇床中培养至OD600为0.4。5 000 r/min条件下离心15 min集菌,用重悬液(10 mmol/L MgCl2、10 mmol/L 2-(N-吗啉代)乙磺酸(pH=5.6)、100 μmol/L乙酰丁香酮)重悬菌体,调节OD600为0.4。使用注射器将菌液注射至烟草叶片下表皮,放置于25 ℃培养48 h。用打孔器在烟草叶片注射区避开针孔打取直径为0.5 cm的叶盘,进行GUS染色分析。
1.2.6 GUS组织染色分析
不同苗龄转基因拟南芥的叶、茎、花、根等器官以及不同处理的拟南芥幼苗和烟草叶盘,取样后置于1.5 mL离心管中,加入适量GUS染色液浸没材料。37 ℃培养箱恒温培养过夜,GUS组织化学染色,之后使用50%乙醇浸泡20 min,再依次使用75%、90%乙醇进行脱色。完全脱色的材料放置在显微镜下观察其组织染色情况。
GUS染色液配方为:2 mmol/L K3[Fe(CN)6]、2 mmol/L K4 [Fe (CN)6]、0.1% Triton X-100、20% CH3OH2、10 mmol/L Na2EDTA、50 mmol/L NaH2PO4和Na2HPO4(pH=7.0)、2 mmol/L X-Gluc[34]。配制好的GUS染色液于避光低温环境下保存。
1.2.7 GUS酶活分析
GUS酶活提取液配方:0.5 mL 0.1 mol/L磷酸缓冲液(pH=7.0)、10 μL 10% SDS、20 μL 0.5 mol/L EDTA(pH=8.0)、1 μL Triton X-100、1 μL β-巯基乙醇,加水定容至1 mL。
取对照、激素和非生物胁迫处理后的适量烟草叶盘,液氮研磨,加入1 mL配制好的酶活提取液,冰上充分混合,将混合液放置于离心管中,4 ℃、12 000 r/min离心5 min,上清液移至新的1.5 mL离心管中,-80 ℃保存。
通过考马斯亮蓝法测定样本的蛋白浓度[35]。GUS酶活测定参考陈婷婷等的方法[36]。将20 μL GUS蛋白提取上清液用无菌水定容至1 mL,加入3 mL G-250,室温反应30 min,使用分光光度计测量595 nm波长处的吸光度,标准曲线读数获得蛋白含量。配制4-MU 1 mmol/L母液(使用终止液定容到250 mL),并配制其梯度浓度液,使用分光光度计在激射光365 nm、发射光455 nm、狭缝10 nm的条件下,测量5种样品的荧光值,并绘制曲线。将配制好的反应缓冲液置于37 ℃预热备用,取10 μL GUS蛋白提取上清液,加入390 μL预热反应缓冲液,立刻取100 μL加入900 μL的反应终止液,37 ℃温浴;之后分别在不同时间段各取100 μL反应液,分别加入900 μL反应终止液。测量各样品的荧光值,绘制测定曲线。使用测定曲线与蛋白含量计算5种样品的酶活性。
2 结果与分析
2.1 VvAGL12启动子克隆
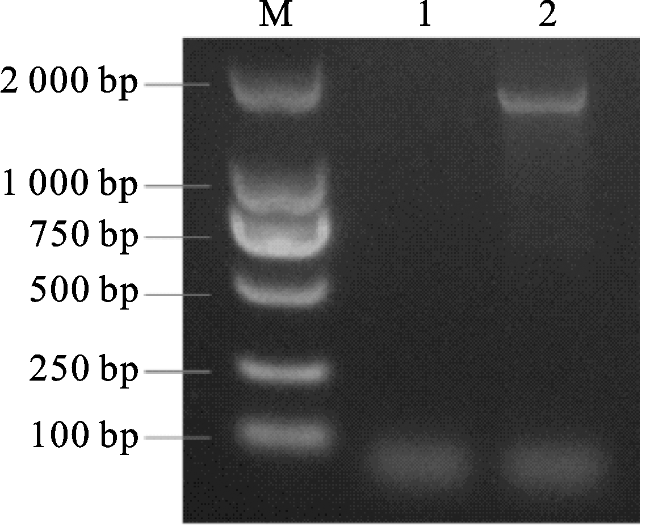
以葡萄基因组DNA为模板,使用特异性引物进行PCR扩增,得到目的条带约1.8 kb。将片段进行胶回收,再与pCAMBIA1300+pBI101载体连接,连接产物转化大肠杆菌DH5α后进行菌落PCR验证(图1)。验证的菌液提取质粒并进行双酶切验证,酶切正确的菌液测序获得启动子序列,序列长度为1 767 bp。
2.2 VvAGL12启动子序列分析
通过PlantCARE和PLACE数据库对proVvAGL12启动子序列进行分析,发现proVvAGL12启动子的响应元件不仅有常见的决定转录起始与转录效率的TATA-box、CAAT-box和光响应元件(G-Box、g-Box、CAG-motif、GATA-motif),在非生物胁迫方面还包括与干旱相关的MYB、MYC响应元件及与低温相关的LTR响应元件;同时,也包括赤霉素(GA3)响应元件(P-box)、ABA响应元件(ABRE)、生长素响应元件(TGA-element)、水杨酸响应元件(TCA-element、W box)、茉莉酸甲酯响应元件(CGTCA-motif、TGACG-motif)以及其他作用元件(图2,表1)。
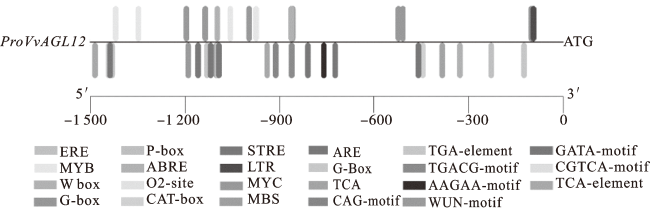
表1 ProVvAGL12启动子顺式作用元件序列及功能Tab.1 Sequences and functions of cis-acting elements on proVvAGL12 promoter |
| 顺式作用元件 | 序列 | 功能 | 数量 |
|---|---|---|---|
| ABRE | ACGTG | ABA响应元件 | 2 |
| ARE | AAACCA | 厌氧诱导响应元件 | 2 |
| AT-rich element | ATAGAAATCAA | AT-rich DNA结合蛋白(ATBP-1)的结合位点 | 2 |
| CAG-motif | GAAAGGCAGAC | 光响应元件 | 1 |
| GATA-motif | AAGGATAAGG | 光响应元件 | 1 |
| G-Box | CACGTT | 光响应元件 | 1 |
| G-box | TAACACGTAG/CACGTC | 光响应元件 | 2 |
| CAT-box | GCCACT | 分生组织调控元件 | 2 |
| CGTCA-motif | CGTCA | 茉莉酸甲酯响应元件 | 1 |
| TGACG-motif | TGACG | 茉莉酸甲酯响应元件 | 1 |
| LTR | CCGAAA | 低温响应元件 | 1 |
| MBS | CAACTG | MYB干旱诱导响应元件 | 1 |
| O2-site | GTTGACGTGA | 玉米代谢调节响应元件 | 2 |
| P-box | CCTTTTG | 赤霉素响应元件 | 1 |
| TCA-element | CCATCTTTTT | 水杨酸响应元件 | 2 |
| W box | TTGACC | 水杨酸响应元件 | 1 |
| TGA-element | AACGAC | 生长素响应元件 | 2 |
| MYB | WAACCA | 脱水响应元件 | 6 |
| MYC | CANNTG | 脱水及低温响应元件 | 7 |
| CAAT-box | CAAT | 启动子和增强子区调控元件 | 25 |
| AT~TATA-box | TATATA | 启动子保守序列 | 13 |
注:序列中W代表T/A,N代表A/T/G/C。 |
2.3 ProVvAGL12转基因株系验证
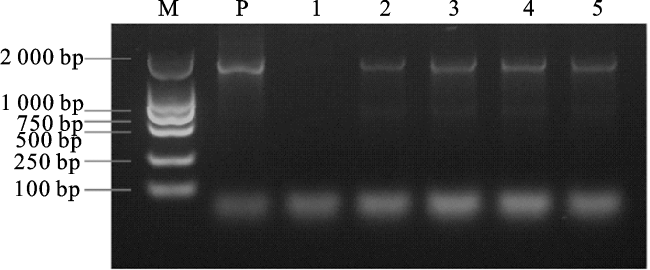
提取野生型拟南芥(Col-0)和proVvAGL12转基因拟南芥植株DNA为模板,用基因克隆引物proVvAGL12-F/R进行PCR验证。以质粒为模板作为阳性对照,以野生型拟南芥DNA为模板作为阴性对照,以proVvAGL12转基因不同株系DNA为模板进行PCR验证。PCR结果显示,阴性对照未出条带,以proVvAGL12转基因植株DNA为模板的PCR条带与阳性对照条带基本一致,表明P1300-proVvAGL12载体成功转入拟南芥基因组(图3)。
2.4 ProVvAGL12驱动GUS在拟南芥多个组织中表达
为进一步研究VvAGL12启动子的调控作用与组织化学定位,将构建好的重组质粒转入农杆菌GV3101,通过花浸法得到转基因拟南芥株系,后续筛选出T3代纯合转基因proVvAGL12拟南芥种子。

综上,proVvAGL12启动子片段在拟南芥中具有启动活性,其驱动GUS在转基因拟南芥植株的叶片、茎段、花器官、根、果荚部位表达,且随着时间的变化不同组织的表达活性有差异,其驱动的GUS表达活性可持续整个生长周期。
2.5 proVvAGL12在拟南芥中的活性分析
2.6 proVvAGL12在烟草中的瞬时表达活性分析
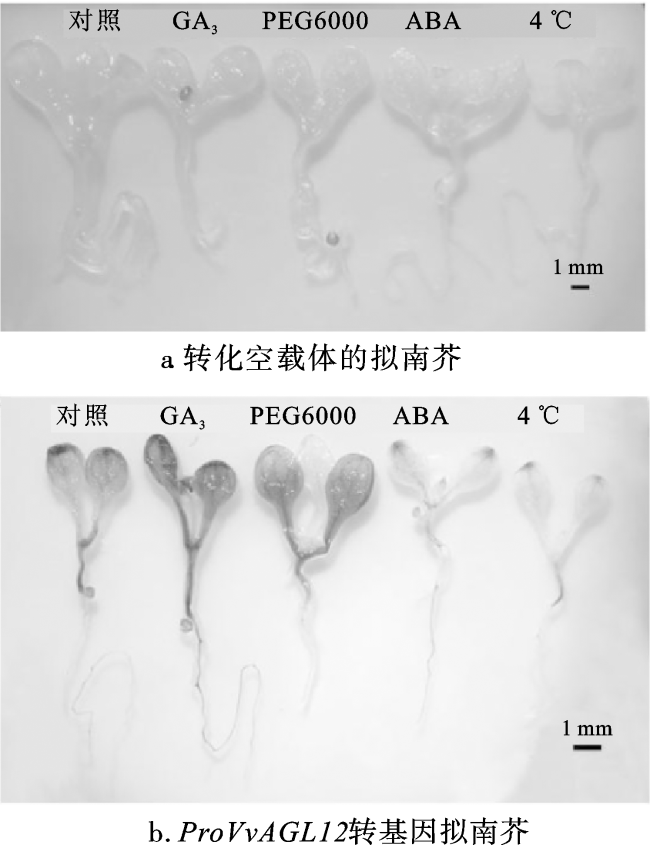
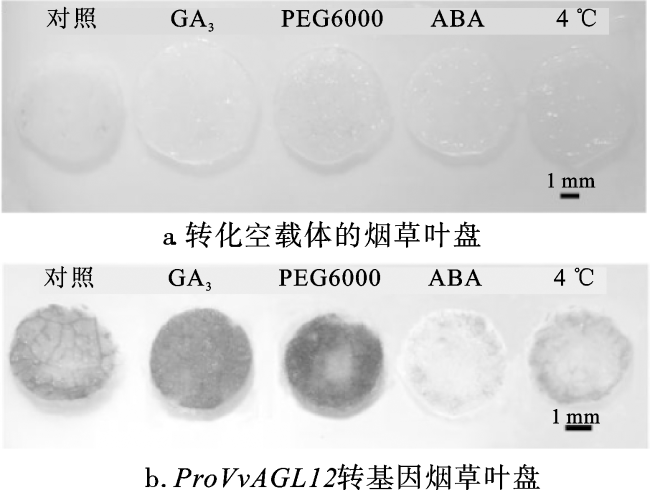
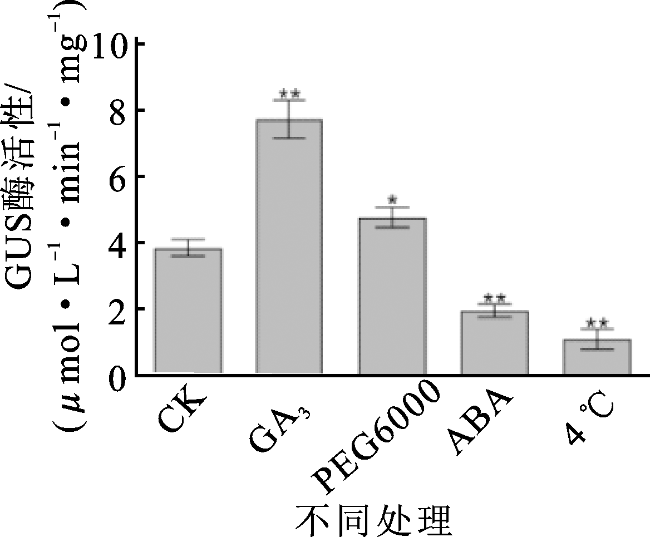
进一步探究proVvAGL12对激素和非生物胁迫的响应情况,在本氏烟草叶片中瞬时表达pCAMBIA1300+pBI101空载体和P1300-proVvAGL12,并进行GUS组织化学染色和定量分析。烟草叶盘GUS染色结果显示,瞬时转化空载体的叶盘在各种处理下均无GUS表达活性;瞬时转化P1300-proVvAGL12的叶盘在5种处理条件下均有GUS表达活性。与对照组相比,GA3与PEG6000处理显著增加了GUS表达活性,而4 ℃与ABA处理显著抑制烟草细胞的GUS表达活性(图6)。同时,将GA3、PEG6000、ABA、4 ℃处理后的烟草叶盘进行GUS酶活性定量分析,发现与对照组相比,GA3、PEG6000处理后的P1300-proVvAGL12转基因叶盘GUS酶活性显著增加,而ABA、4 ℃处理的转基因叶盘GUS酶活性显著降低(图7)。综上所述,proVvAGL12在烟草中具有启动活性,且对GA3、PEG6000、ABA、4 ℃处理产生响应,但不同处理的响应方式有所不同。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 讨论
3.1 MADS-box家族基因启动子研究
苹果中,外源喷施赤霉素显著诱导MdMADS50基因的表达,后续研究发现赤霉素诱导MdMADS50启动子的表达活性增加[40]。玉米中,启动子的表达活性研究促进了对玉米MADS-box家族基因功能的理解[41⇓-43]。沙荆中,MADS-box家族基因启动子的光响应元件及脱落酸、赤霉素、低温、防御相关的响应元件数目相对较多,可能在参与植物生长发育过程中起着重要作用[44]。米槠中,分析启动子顺式作用元件发现,逆境胁迫元件与植物生长发育相关元件较多,米槠MADS-box在响应生物胁迫、非生物胁迫、激素诱导、植株生长发育等方面具有重要作用[45]。葡萄中关于MADS-box家族启动子的调控活性未得到广泛研究,进一步发掘proVvAGL12的表达趋势、顺式作用元件及胁迫响应方式,可为研究VvAGL12的激素与非生物胁迫调控以及具体基因功能提供参考。
3.2 GUS活性受GA3、ABA、PEG和低温的影响
GA3是最早被发现并使用的激素之一,根据其结构合成的化学物质是常见的诱导葡萄无核果实和膨大果实的生长调节剂[44]。GA3不仅在植物生长发育方面起重要调控作用,在响应非生物胁迫方面也发挥着重要作用[46-47]。已有研究发现,VvAGL11与VvAGL15基因的表达水平受到GA3的抑制,通过响应GA3影响种胚发育、抑制胚形成可达到葡萄无核果实生长的目的[48]。GA3处理植物后,通过提高植物SOD、POD、CAT等抗氧化酶的活性,改变渗透调节物质以及相关激素的含量,来提高植物的抗盐性[49]。ABA作为常见的植物内源激素,在抑制植物生长发育和参与逆境胁迫应答方面发挥着重要作用[50-51]。经过一定浓度的外源ABA处理或内源ABA积累后,可调节植物气孔运动,改变植物水分利用率,达到植物抗旱、抗寒的作用[52-53]。
从葡萄基因组中将MADS-box家族VvAGL12基因上游1 767 bp的启动子序列提取分离出来,并对该序列进行预测和功能验证,分析proVvAGL12驱动的GUS在拟南芥中的组织表达模式,探讨激素和非生物胁迫对GUS的调控作用。研究发现拟南芥幼苗叶部的维管组织GUS表达活性强于根部的维管组织,但随着植株生长,二者GUS活性差距变小,同时发现营养阶段的GUS活性要强于生殖阶段(图4),说明proVvAGL12驱动的GUS表达具有时空特性。
本研究发现VvAGL12启动子区域含有多个干旱响应元件、低温响应元件、ABA响应元件和赤霉素响应元件,推测VvAGL12的表达受到ABA、GA3、低温和干旱的调控。为了进一步验证,将proVvAGL12序列分别在拟南芥和烟草中进行异源转化,并对转基因植株分别进行GA3、ABA、PEG和低温处理。研究发现与对照相比,GA3处理显著诱导了proVvAGL12驱动的GUS活性表达,推测该启动子在葡萄抗盐胁迫与果实无核生长发育方面具有调控作用,为培育优质抗逆无核葡萄品种奠定了基础。结合顺式作用元件分析与实验数据发现,经过PEG处理的转基因植株,GUS活性受到PEG诱导且其表达活性明显增强;而经过ABA与低温处理后的转基因植株,GUS表达活性显著减弱,其响应模式与GA3、PEG不同。综上,proVvAGL12驱动的GUS活性受GA3、ABA、PEG和低温调控,推测VvAGL12是葡萄响应干旱胁迫、盐胁迫、低温逆境胁迫的相关基因,将该基因转入葡萄与其他植物中或能提高植物的抗旱、抗盐、抗寒能力,减少季节性与环境对植物的影响,对植物培育和抗逆生长有参考价值,而VvAGL12是否受激素与非生物胁迫调控及其具体基因功能还有待验证。
4 结论
proVvAGL12驱动的GUS活性受GA3、ABA、PEG和低温调控。经低温和ABA处理后GUS表达活性与对照相比显著降低,经PEG6000和GA3处理后GUS表达活性则显著升高,说明低温和ABA对proVvAGL12驱动的GUS表达活性具有抑制作用,而GA3和PEG6000则有促进作用。ProVvAGL12驱动的GUS活性对激素与非生物胁迫都存在响应但响应模式不同,可将其运用在以后的葡萄育种生产中,对葡萄的抗盐、抗旱、抗寒、实现果实无核提供理论基础。