

随着现代工业的发展和城镇化进程的不断推进,重金属镉(cadmium,Cd)的污染日益严重,镉及镉化合物已被世界卫生组织国际癌症研究机构列入有毒有害水污染物名录和致癌物清单。一般来讲,植物能够耐受低浓度的镉,但镉过量会使植物中毒,严重威胁作物安全[1]。镉污染对植物的伤害是多方面的,其在植物不同部位的吸收和积累程度不同,毒副作用也不同。当镉在植物中积累到一定程度时,植物会出现中毒症状,叶片会变黄并出现斑点,致使植物生长发育缓慢甚至死亡[2]。镉影响植物的生命活动主要体现在光合作用、呼吸作用等方面[3-4]。镉被植物吸收后在细胞中积累,阻碍叶绿体发育,影响光合色素合成,进而抑制植物的光合作用[5-6]。镉还会通过离子通道进入保卫细胞,致使气孔关闭,呼吸作用受到阻碍,植物需要调节呼吸作用来制造能量,产能负荷加大,进而抑制蒸腾作用,造成植物早衰[6⇓-8]。此外,镉极易被植物吸收并通过食物链转移到动物和人身上,危害人和动物的健康[9⇓-11]。
重金属相关异戊二烯化植物蛋白(heavy metal-associated isoprene plant protein,HIPP)是维管植物中一种独特的与金属结合的金属伴侣蛋白,其具有1个重金属相关结构域和1个异戊二烯结构域[12-13],主要参与重金属稳态反应以及解毒、耐受性等响应机制[14],对重金属镉的反应尤为明显[15]。研究发现玉米中应答镉胁迫的保守基因包含3个HIPP,分别为ZmHIPP27、ZmHIPP35和ZmHIPP36,三者在玉米应对非生物胁迫中都发挥了重要作用[16]。对水稻转录组进行分析,发现HIPP基因的表达在镉胁迫下发生改变,其中,OsHIPP42被认为是参与镉胁迫反应的关键基因[12],OsHIPP16在酵母突变体ycf1中的异位表达提高了酵母对镉和锌的耐受性[17]。超累积器植物圆锥南芥HIPP26基因在拟南芥中异位表达可以增强植株对镉的耐受性,同时增加镉在植株中的累积[18]。
小麦(Triticum aestivum)是世界上最重要的粮食作物之一,在解决世界饥饿问题和确保全球粮食安全方面具有重要作用。粗山羊草(Aegilops tauschii)是普通小麦D基因组的供体,具有多种抵御生物和非生物胁迫基因,是改良小麦的潜在遗传资源[19-20]。尽管在植物中发现HIPP20、HIPP21、HIPP22、HIPP26、HIPP27等基因可通过结合镉而发挥解毒功能[21],但粗山羊草中有关HIPP基因的研究仍然有限。
本课题组前期利用RNA测序分析了粗山羊草在镉胁迫下的差异表达基因,发现AetHIPP28基因在镉胁迫下表达上调[22]。本研究通过农杆菌介导获得过表达AetHIPP28的转基因拟南芥,并分析转基因拟南芥在镉胁迫下的生理生化指标以及Cd2+在根部的内流和分布,探究AetHIPP28响应镉胁迫的作用机制。
1 材料和方法
1.1 植物材料及生长条件
粗山羊草(种质编号KU2126)种子用5%的NaClO溶液浸润,倒置并震荡 8 min,使用无菌水洗净表面残余溶液。种子放置在培养皿中,置于25 ℃的恒温培养箱内,按16 h/8 h的光暗周期进行培养。将已发芽的种子转移到Hoagland营养液中(1/2标准浓度),营养液每天更换。待植株生长至三叶期时进行Cd处理,处理组为Hoagland营养液(1/2标准浓度)+50 μmol/L CdCl2,对照组为Hoagland营养液(1/2标准浓度)。分别在处理3 h和7 d后取样,进行基因表达分析并检测植株的生理生化指标。
1.2 AetHIPP28基因的克隆
使用EASY spin plus试剂盒(Aidlba Biotech,中国)提取粗山羊草总RNA。使用HiFiScript gDNA Removal cDNA Synthesis Kit(CwBio,中国)合成第一链cDNA。使用PCR仪(Eastwin,中国)进行目的基因扩增。PCR产物通过1%琼脂糖凝胶电泳检测,并使用凝胶提取试剂盒(CwBio,中国)进行纯化。纯化产物与pMD18-T载体(Takara,中国)连接,并转化到DH5α细胞中进行测序。测序由生工生物工程(上海)股份有限公司完成。
1.3 逆转录实时定量聚合酶链式反应(reverse transcription real-time quantitative polymerase chain reaction,RT-qPCR)
为检测AetHIPP28基因对Cd胁迫的响应,利用EASY spin plus试剂盒(Aidlba Biotech,中国)分别从粗山羊草的嫩叶和根部提取总RNA,经反转录合成cDNA(CwBio,中国)。RT-qPCR反应在Applied Biosystems QuantStudio 3 Real-Time PCR系统(ThermoFisher,美国)上进行,使用PowerUpTM SYBRTM Green mater Mix(Bio-Rad, 美国)进行RT-qPCR分析。如Livak等[23]所述,使用2-ΔΔCt方法计算基因的相对表达水平。
1.4 亚细胞定位
将删除终止密码子的AetHIPP28编码区连接到pROKⅡ-GFP载体上,通过农杆菌介导转化,将重组质粒注射到烟草(Nicotiana benthamiana)叶片中,利用pROKⅡ-GFP作为对照。暗培养3 d后,使用荧光显微镜(Nikon,日本)观察AetHIPP28-GFP的荧光信号。
1.5 AetHIPP28在拟南芥中的遗传转化与鉴定
野生型(wild type,WT)和转基因株系在MS培养基(海博生物,中国)中生长3周,然后转移到含有50 μmol/L Cd(以化合物CdCl2·5H2O计)的营养土中生长2周。使用GUS染色试剂盒(MeilunBio,中国)检测WT和转基因株系的GUS活性。
1.6 生理生化指标测定
采集对照和50 μmol/L Cd处理7 d的3周龄拟南芥,使用检测试剂盒(Solarbio,中国)测量WT和转基因拟南芥组织中的超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)、过氧化氢酶(catalase,CAT)的活性以及脯氨酸、叶绿素、丙二醛(malondialdehyde,MDA)、谷胱甘肽(glutathione,GSH)的含量。参照Zhang等[26]所述的电感耦合等离子体质谱法测量WT和转基因拟南芥根分生组织中的Cd2+净通量。使用荧光探针LeadmiumGreen AM(Invitrogen,美国)检测根组织中的Cd分布[27]。
1.7 数据分析
利用SPSS 13.0软件对实验数据进行统计学分析,采用ANOVA进行单因素方差分析。所有数据至少根据3个独立生物学重复得出。
1.8 引物
本研究中使用的所有引物信息如表1所示。
表1 研究中使用的引物信息Tab.1 Primer information used in this study |
| 引物 | 序列(5'-3') | 用途 |
|---|---|---|
| F1 | ACGCACACCGCAACTACAACAG | AetHIPP28的qPCR检测 |
| R1 | CGTGGAGGTCCGAGTCGTAGTAG | |
| F2 | AAGATGGATTGCACGCAGGT | 检测表达AetHIPP28的转基因拟南芥 |
| R2 | TCACGGGTAGCCAACGCT | |
| F3 | GCTCTAGAATGACGATCGTGGAGATG | 克隆AetHIPP28基因的开放阅读框 |
| R3 | CGGGATCCCATGACGGAGCAGGCG | |
| F4 | TGAGACTTTTCAACAAAGGGTAATA | 跨载体检测转基因拟南芥 |
| R4 | TGGTAGGAGTAATAGGCCGGCTGT | |
| F5 | GCTGGCTCGTTCAACTGATG | 26S内参基因 |
| R5 | GGACCAAGCGTTCTGATTACTC |
注:F3的Xba Ⅰ和R3的BamH Ⅰ的限制性位点加粗表示。 |
2 结果
2.1 AetHIPP28亚细胞定位
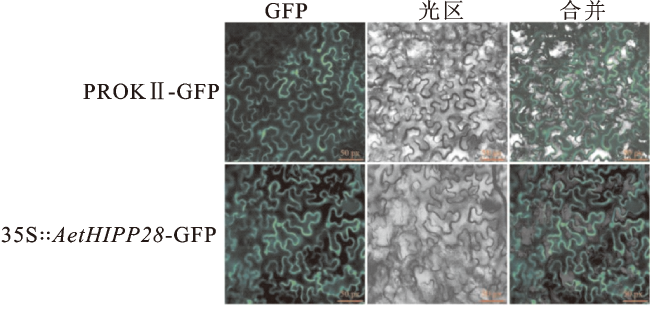
将35S∶∶AetHIPP28-GFP融合载体注射烟草表皮细胞,以pROK Ⅱ空载体注射烟草表皮细胞作为对照。AetHIPP28-GFP在核膜和质膜都有明显荧光(图1),表明AetHIPP28在细胞核和质膜中均能表达。
2.2 Cd对AetHIPP28在粗山羊草中表达的影响
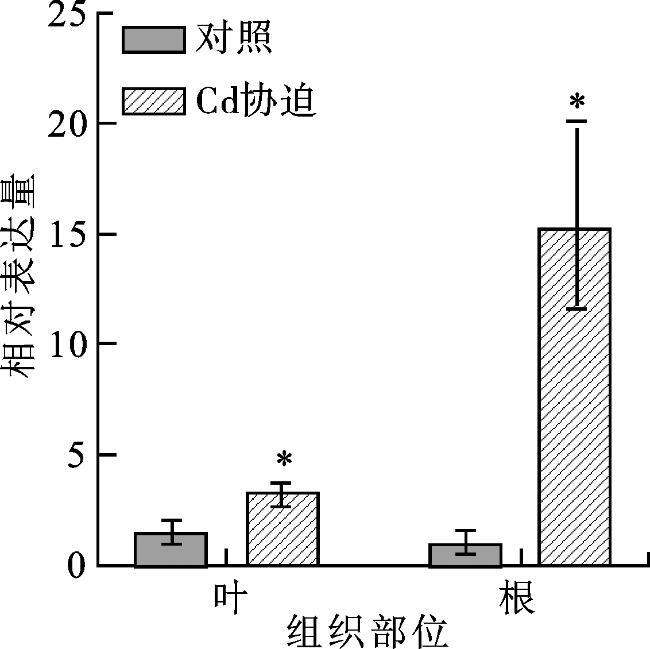
通过RT-qPCR分析Cd胁迫下粗山羊草中AetHIPP28基因在根和叶中的表达情况。结果表明,在Cd的胁迫下AetHIPP28基因在根和叶中均上调表达,在根部诱导上调的表达量高于叶中,表明AetHIPP28主要在粗山羊草的根部响应Cd胁迫(图2)。
2.3 转基因拟南芥的鉴定
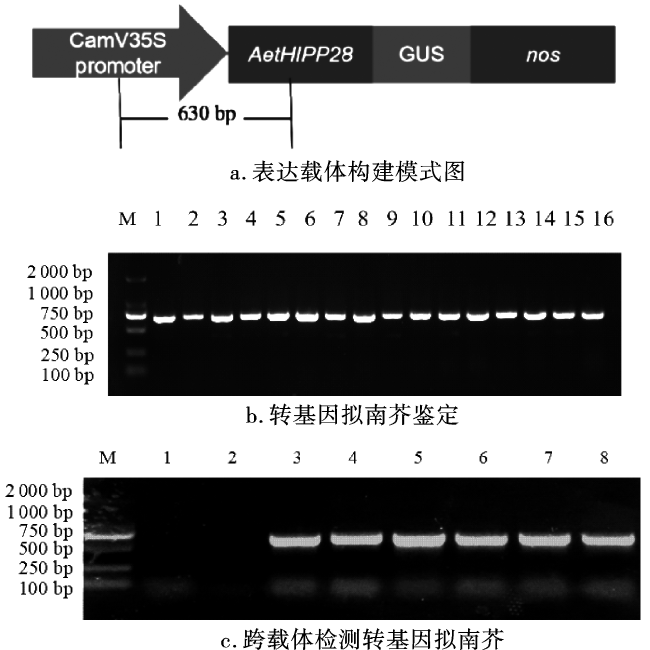
为初步确定AetHIPP28基因在转基因拟南芥中的表达,对转基因拟南芥进行GUS染色分析,通过GUS组织染色排除假阳性拟南芥。与WT相比,转基因拟南芥的叶片和根部均能启动GUS表达(图4),说明AetHIPP28基因可以在转基因株系中表达。未经Cd处理时,WT拟南芥整株呈现浅绿色,转基因株系的部分叶子和根呈浅蓝色。经Cd处理后,WT拟南芥仍然保持浅绿色,而转基因株系的颜色显示为深蓝色。组织化学染色结果表明,Cd处理后AetHIPP28-GUS融合基因的表达量明显增加。

2.4 Cd胁迫下转基因拟南芥的表型分析
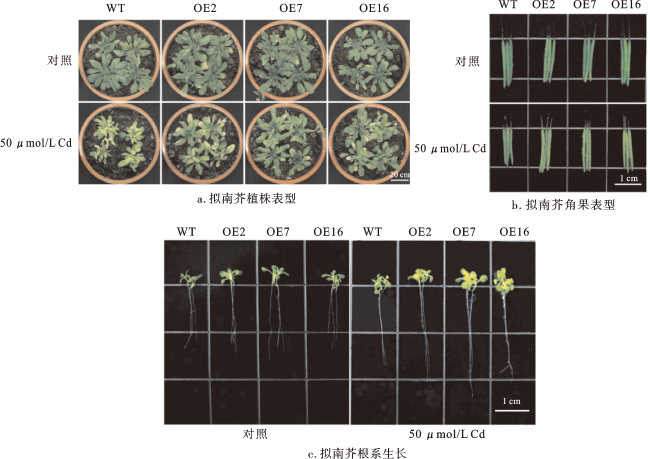
通过将AetHIPP28的ORF引入pBI121载体产生1个重组质粒,并将其转入拟南芥,表达AetHIPP28的转基因株系通过PCR进行鉴定,以阐明AetHIPP28在Cd胁迫下的功能。随机挑选3个转基因拟南芥株系OE2、OE7和OE16进行后续研究。
2.5 AetHIPP28对转基因拟南芥生理参数的影响
对WT和转基因拟南芥在Cd处理前后的SOD、POD、CAT活性进行分析,结果表明(图6A~C),在对照组中,WT和转基因株系之间的SOD、POD、CAT活性没有显著差异,但Cd处理提高了转基因株系和WT植株中抗氧化剂的活性,且转基因株系中SOD、POD、CAT活性的增加幅度明显高于WT植株。
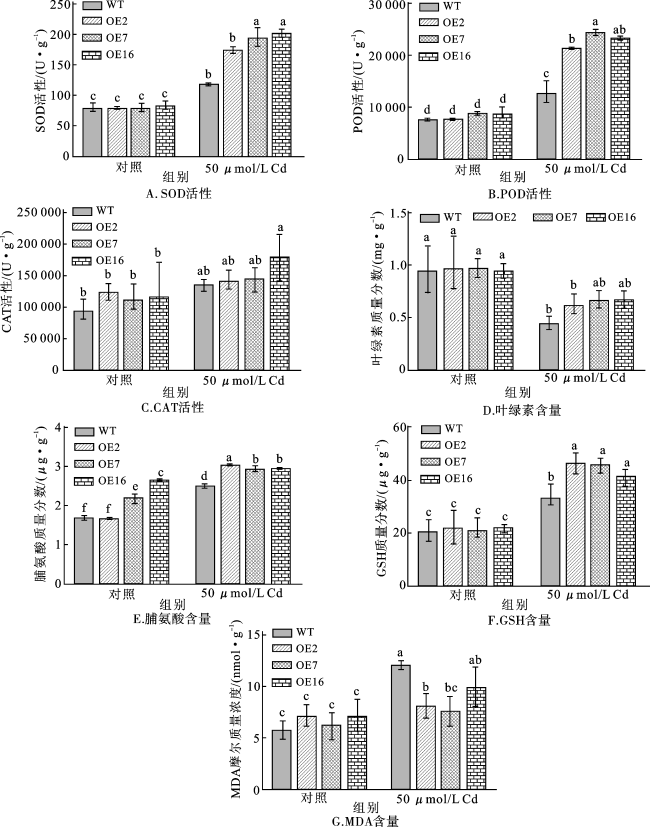
进一步比较WT和转基因株系在Cd胁迫前后叶绿素、脯氨酸和GSH的含量,结果显示(图6D~F),WT植株的叶绿素含量在Cd胁迫下受到明显抑制,但是转基因株系中除OE2外,其他植株变化不明显。在没有Cd处理的情况下,过表达株OE2与WT的脯氨酸含量无显著差异,但在过表达株OE7和OE16中脯氨酸含量显著提高。对照组中转基因拟南芥和WT之间的GSH含量没有显著差异。值得注意的是,经Cd处理后,转基因拟南芥中脯氨酸和GSH的积累与WT相比均明显增加。这些结果说明,过量表达AetHIPP28基因使植株积累了更多的脯氨酸和GSH,从而提高了植株对Cd胁迫的耐受性。
由图6G可以看出,正常条件下生长的幼苗中MDA含量较低,不同株系之间没有明显差异。经Cd处理后,拟南芥中的MDA含量显著增加,但转基因拟南芥中的MDA含量比WT植株中低。这说明在Cd胁迫下,WT植株的细胞膜损伤更为严重,进一步表明AetHIPP28的表达可以减少植物细胞膜的损伤,从而提高植株对Cd胁迫的耐受性。
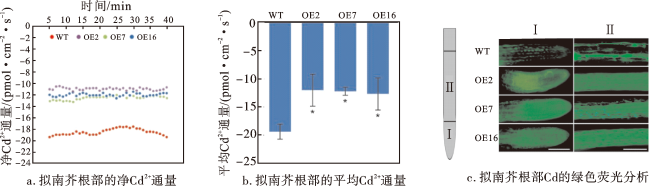
2.6 过表达AetHIPP28对拟南芥Cd2+内流和分布的影响
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 讨论
先前研究已充分证明,HIPP基因家族在植物应对重金属胁迫过程中具有关键作用,包括重金属稳态和解毒等过程。本研究结果进一步证实,AetHIPP28基因能够提高植物的耐Cd性,并减轻Cd对植物根系发育造成的损伤。Khan等[12]在水稻全基因组中共发现了54个HPP和HIPP基因,其中有11个HIPP基因在Cd的胁迫下上调。此外,研究发现OsHIPP42突变株在Cd处理下的生长更弱,表明HIPP基因能够降低重金属对植物的毒害作用。Zhang等[11]研究发现,HvHIPP1基因可以增强小麦对Cd的耐受性。本课题组基于先前的转录组数据,发现粗山羊草HIPP28基因受Cd诱导上调[22]。本研究通过RT-qPCR发现AetHIPP28在粗山羊草的根部响应Cd胁迫更为明显,这与转录组结果是一致的,由此推测,AetHIPP28基因可能在粗山羊草根部对Cd元素的积累和超耐受过程发挥关键作用。为进一步探究AetHIPP28基因的功能,本研究通过拟南芥的遗传转化,对AetHIPP28转基因拟南芥进行耐Cd性分析,并开展AetHIPP28功能验证。
在重金属胁迫下,植物体内活性氧(reactive oxygen species,ROS)代谢受阻,ROS过量积累,造成膜脂质过氧化并破坏电子传递链,从而导致植物氧化损伤[36]。活性氧清除能力是衡量植物抗逆性的重要指标,植物SOD、POD和CAT是保护性酶,可以清除活性氧,协助植物抵抗胁迫[37]。本研究通过测定抗氧化酶活性发现经Cd处理后,野生型拟南芥的酶活性显著低于转AetHIPP28拟南芥,表明AetHIPP28基因可能参与调控SOD、CAT、POD等酶活力,从而加强植物对ROS的清除能力,提高植物对非生物胁迫的耐受性。逆境会引起植物细胞渗透胁迫,从而影响植物生长。MDA和脯氨酸是植物调节细胞渗透压的重要物质,植物在遭受迫害时,体内会积累大量的MDA和脯氨酸[38-39],因此MDA和脯氨酸可以作为抗逆育种的生理指标。经Cd处理后,WT拟南芥脯氨酸的积累量显著高于AetHIPP28转基因拟南芥,而WT拟南芥的MDA积累量显著低于AetHIPP28转基因拟南芥。以上结果说明AetHIPP28基因能增强植物对ROS的清除能力,减少ROS积累,并能够提高脯氨酸积累,从而参与植物的渗透调节,提高植物对Cd的耐受性。
降低Cd2+的跨膜运动对植物耐Cd性具有重要意义。本研究采用电感耦合等离子体质谱法检测了根尖表皮细胞对Cd2+的反应,结果表明过表达AetHIPP28基因改变了植株的实时Cd2+通量,Cd2+内流被阻断,说明过表达AetHIPP28能够显著抑制Cd2+内流。这种阻断能力可能与HIPP蛋白的Cd结合能力有关,类似金属螯合剂或者金属转运蛋白阻断Cd2+内流的机制[40]。同时,转基因拟南芥的根部也增加了Cd累积,推测过表达AetHIPP28不仅能够增强植物的耐Cd性,还可以减少Cd在植物体中的向上转运。这也进一步解释了在粗山羊草受Cd胁迫后AetHIPP28基因在根部表达量显著提高的现象。
4 结论
AetHIPP28蛋白定位于质膜和细胞核中,AetHIPP28主要在粗山羊草的根部响应Cd胁迫,过表达AetHIPP28通过调控抗氧化酶活性提高转基因拟南芥的耐Cd性,同时抑制Cd2+的内流,并增强根部Cd的积累。本研究明确了AetHIPP28基因耐Cd胁迫的功能,为解析其耐Cd分子机制和利用该基因创制耐Cd植物奠定了基础。