

糖尿病肾病(diabetic nephropathy,DN)是由微血管病变引起的主要糖尿病并发症,也是糖尿病致死的主要原因之一[1-2]。糖尿病肾病的发生受多种因素影响,其病理生理学非常复杂,包括遗传因素、血流动力学的改变、氧化应激、炎症和细胞因子的作用以及糖代谢等方面。高血糖通过影响多种代谢和信号通路,成为引发肾损伤的关键因素之一;当体内糖代谢过程发生紊乱时,易引发糖尿病肾病[3]。上皮间充质转化(epithelial-mesenchymal transition,EMT)是上皮细胞按照特定程序获得间充质表型的一种生物学现象[4-5],它参与多种生理和病理过程,发生在胚胎发育、炎症、组织纤维化和癌症转移等过程中[6-7]。随着上皮细胞极性的失去,其与邻近细胞和基底膜的连接减少,上皮细胞变得更具侵袭性和迁移性。研究发现,上皮间充质转化的发生与多种标志蛋白表达的改变有关。例如,E-cadherin缺乏是上皮间充质转化过程中的一个关键事件,它与细胞间连接、细胞极性的变化以及细胞外基质的产生有关[8]。在细胞纤维化过程中,平滑肌肌动蛋白的表达大幅升高,这是维持成纤维细胞间充质表型所必需的[9]。此外,上皮间充质转化受多种基因的影响,包括蜗牛蛋白(Snail)和E盒锌指蛋白(Zeb)[10]。足细胞又称作肾小球上皮细胞,是维持肾小球正常滤过功能的细胞之一,高糖等刺激会诱导足细胞发生上皮间充质转化,从而影响肾脏的过滤功能,对蛋白尿以及糖尿病肾病等疾病的发生具有重要影响。高血糖可触发肾小管上皮细胞的上皮间充质转化,导致炎症和纤维化加速[11]。持续的高糖环境可触发和激活多种信号通路[12],使细胞内活性氧增加,细胞间纤维化加剧。目前,发生上述情况的确切生物学过程尚不清楚[13-14]。
泛素化是蛋白质的一种重要翻译后修饰,它调节一系列生理和病理过程,包括细胞生长和迁移、应激反应、癌症等[15]。文献报道,在糖尿病肾病患者中,有大量K63泛素化蛋白聚集,加剧了肾脏损伤和上皮间充质转化[16]。在Ⅱ型糖尿病患者的尿液中发现了泛素蛋白融合产物UbA52,证实了泛素化途径在肾损伤中的关键作用[17]。E2泛素结合酶变异体1(ubiquitin binding enzyme E2 isomer-1, UBE2V1)是UBC13的辅助因子,属E2蛋白家族成员,是一类保留了核心成分但活性中心缺乏半胱氨酸残基的酶,也是已知的唯一一类与K63泛素化途径相关的E2酶[18]。它位于20号染色体,两侧存在多种与Ⅱ型糖尿病相关的基因,研究表明其可能是一个癌基因或肿瘤抑制基因[19-20]。UBE2V1通过抑制自噬可诱导结直肠癌细胞的上皮间充质转化和转移[21];与正常肾脏相比,糖尿病肾病患者的肾脏中UBE2V1的表达量增加,对肾小管间质纤维化和糖尿病肾病的发病具有重要影响[22]。
UHRF家族成员包括泛素样含PHD和环指域蛋白2(ubiquitin-like with PHD and ring finger domains 2, UHRF2)与泛素样含PHD和环指域蛋白1(ubiquitin-like with PHD and ring finger domains 1, UHRF1),UHRF2与UHRF1为同源物,与UHRF1共享一个结构域[23]。研究表明,UHRF2能够调控多个上皮间充质转化相关基因的表达,从而驱动细胞侵袭[24],但UHRF2在糖尿病肾病中的作用尚未研究。有文献称UHRF2在K63泛素化通路中作为E3泛素连接酶起作用,但确切机制仍不清楚。
本研究以人肾小管上皮细胞(human renal tubular epithelial cells,HKC)为模型,探讨UBE2V1和UHRF2在高糖诱导肾小管上皮细胞上皮间充质转化过程中的作用,以期加深对糖尿病肾病发病机制的了解,为糖尿病的治疗提供新思路。
1 材料与方法
1.1 细胞培养
肾小管上皮细胞分别用完全培养基(RPMI 1640培养基+10%胎牛血清+1%青霉素-链霉素)、高糖培养基(完全培养基+30 mmol/L葡萄糖)和甘露醇培养基(完全培养基+30 mmol/L甘露醇)进行培养。
1.2 实时荧光定量
将人肾小管上皮细胞重悬于TRIzol试剂(赛默飞世尔科技有限公司)中,根据生产商说明使用HiScriptⅢ RT SuperMix for qPCR(南京诺唯赞生物科技股份有限公司)合成cDNA,使用 ChamQTM Universal SYBRqPCR Master Mix(南京诺唯赞生物科技股份有限公司)进行定量 RT-PCR。以肌动蛋白为内参,对目的基因的转录进行定量分析。所用引物信息如表1所示。
表1 实验引物信息Tab.1 Experimental primer information |
| 引物 | 引物序列 | 用途 |
|---|---|---|
| actin-F | ACCTTCTACAATGAGCTGCG | qPCR |
| actin-R | CCTGGATAGCAACGTACATGG | |
| UBE2V1-F | TGGAGTGGTGGACCCAAGA | qPCR |
| UBE2V1-R | TAACACTGTCCTTCGGGCG | |
| UHRF2-F | ACCTTCCAATCAGCCATCTAC | qPCR |
| UHRF2-R | CTTCAAACCAAGCACCAAGG | |
| actin-F | ACCTTCTACAATGAGCTGCG | qPCR |
| actin-R | CCTGGATAGCAACGTACATGG | |
| UBE2V1-F | TGGAGTGGTGGACCCAAGA | qPCR |
| UBE2V1-R | TAACACTGTCCTTCGGGCG | |
| shUBE2V1-Top1 | GATCCCCAAGAGCCATATCAGTGCTACTTCCT GTCAGATAGCACTGATATGGCTCTTGGTTTTTG | UBE2V1 RNAi (pGreen-UBE2V1-1) |
| shUBE2V1-Bottom1 | AATTCAAAAACCAAGAGCCATATCAGTGCTAT CTGACAGGAAGTAGCACTGATATGGCTCTTGGG | |
| shUBE2V1-Top2 | GATCCATTTCACTGCAAAGGAGTAAACTTCCT GTCAGATTTACTCCTTTGCAGTGAAAT TTTTTG | UBE2V1 RNAi (pGreen-UBE2V1-2) |
| shUBE2V1-Bottom2 | AATTCAAAAAATTTCACTGCAAAGGAGTAAAT CTGACAGGAAGTTTACTCCTTTGCAGTGAAATG | |
| shUBE2V1-Top3 | GATCCACAAAGCAAACTGAGTGATGACTTCCT GTCAGATCATCACTCAGTTTGCTTTGT TTTTTG | UBE2V1 RNAi (pGreen-UBE2V1-3) |
| shUBE2V1-Bottom3 | AATTCAAAAAACAAAGCAAACTGAGTGATGAT CTGACAGGAAGTCATCACTCAGTTTGCTTTGTG | |
| shUHRF2-Top1 | GATCCCGTCTCTTCTTCCATTACAATCTTCCTGTC AGAATTGTAATGGAAGAAGAGACGTTTTTG | UHRF2 RNAi (pGreen-UHRF2-1) |
| shUHRF2-Bottom1 | AATTCAAAAACGTCTCTTCTTCCATTACAATTCTG ACAGGAAGATTGTAATGGAAGAAGAGACGG | |
| shUHRF2-Top2 | GATCCCTGCTGATGAAGACGTTATTTCTTCCTGTC AGAAAATAACGTCTTCATCAGCAG TTTTTG | UHRF2 RNAi (pGreen-UHRF2-2) |
| shUHRF2-Bottom2 | AATTCAAAAACTGCTGATGAAGACGTTATTTTCTG ACAGGAAGAAATAACGTCTTCATCAGCAGG | |
| UHRF2 BamHⅠ NS | TGACGACGATAAGAGCCCGGGCGGAT CCATGTGGATACAGGTTCGC | UHRF2过表达 (pCMV-UHRF2) |
| UHRF2 HindⅢ CAS | CGAGGTCGACGGTATCGATAAGCTTT CATCGTCCTTTGCTGTAG | |
| UBE2V1 SacⅡ NS | AACAAAAGCTGGAGCTCCACCGCGGAT GTACCCTTATGACGTCCCCGATTACGCC | UBE2V1过表达 (pCMV-UBE2V1) |
| UBE2V1 HindⅢ CAS | CGAGGTCGACGGTATCGATAAGC TTTTAATTGCTGTAACACTGTCC |
1.3 基因沉默和过表达
基因沉默根据System Biosciences公司的产品说明书,通过转染2~3个预选目标基因的shRNA载体进行,选择载体为pGreenPuroTMshRNA。基因过表达选择pCMV-tag2B连接基因构建过表达载体,使用Lipofectamine 2000转染试剂(赛默飞世尔科技有限公司)在HKC细胞中转染。
1.4 划痕测试
将人肾小管上皮细胞均匀涂布于六孔培养板中,过夜后采用lip2000转染法将质粒pGreen-UBE2V1、pCMV-UBE2V1、pGreen-UHRF2和pCMV-UHRF2转染到细胞中,待48 h左右细胞完全黏附后进行划痕实验。划痕时弃去培养基,用200 μL移液管吸头沿着六孔板划痕细胞。划痕后,向六孔板中加入适量的无菌磷酸盐缓冲溶液(phosphate buffered saline,PBS),轻轻摇动六孔板以清洗细胞,去除划伤细胞,重复此步骤3次至划伤后留下的缝隙清晰可见。每孔加入2 mL培养基(实验设计中规定的不同无血清培养基),置于37 ℃、5% CO2培养箱中培养。分别于培养0、24、48 h时拍照,使用Image J测量划痕面积。通过比较划痕面积的变化确定伤口愈合率,并进一步比较细胞迁移率,最后使用Graphpad prism 8.0绘图。
1.5 免疫印迹分析
将细胞重悬于含蛋白酶抑制剂的RIPA裂解缓冲液(25 mmol/L Tris-HCl, 150 mmol/L NaCl, 1%脱氧胆酸钠, 0.1%十二烷基磺酸钠, pH 7.4)中,用组织研磨器完全匀浆,7 500 r/min、4 ℃条件下离心20 min去除碎片。用BCA法对样品进行定量,然后将15~30 μg样品加入含5% β-巯基乙醇的十二烷基硫酸钠(sodium dodecyl sulfate,SDS)上样缓冲液中,100 ℃煮沸5 min。将虫体裂解液载入SDS-PAGE并转移到PVDF膜上,用一抗(β-actin负载对照)和HRP结合的二抗探针检测,并用Immobilon Western chemiluminescent HRP底物显色,PVDF膜用MiniChemi610显色。
1.6 K63泛素化蛋白的分离和免疫沉淀
细胞传代后,均匀铺在10 cm的平板上,第2天将质粒pCMV-UHRF2转染到相应的平板上。转染48 h后,转入相应的完全培养基或高糖培养基,加入MG132蛋白酶体抑制剂至终物质的量浓度为5 μmol/L。24 h后收集样品,用抗FLAG M2亲和胶(SIGMA, A2220)免疫沉淀携带FLAG标签的蛋白(4 mg),4 ℃搅拌孵育4~8 h。
1.7 统计分析
使用GraphPad prism 8.0软件分析定量数据;误差条代表平均值的标准误差;两组平均值的比较采用双尾非配对t检验。以*、**和***分别代表P<0.05、P<0.01和P<0.001水平差异显著。
2 结果
2.1 高糖提高UHRF2在HKC中的表达并引发EMT
分别用高糖培养基(30 mmol/L 葡萄糖)和甘露醇培养基(30 mmol/L 甘露醇)处理肾小管上皮细胞1 h、2 h、4 h和8 h,qPCR检测UHRF2的mRNA表达水平,结果如图1a所示。用甘露醇处理细胞时,对照组和处理组UHRF2的mRNA表达水平无明显差异;但用葡萄糖处理时,处理组UHRF2的表达量明显高于对照组,且在处理1 h和2 h时有明显差异。
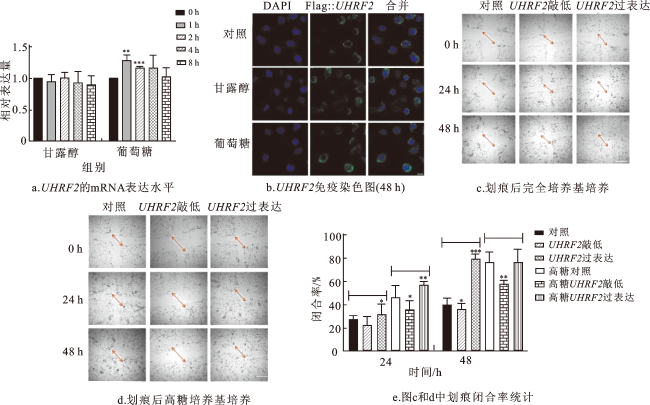
正常情况下,免疫染色显示UHRF2在细胞质中表达(图1b),加入甘露醇和高糖后,UHRF2仍在细胞质中表达。这表明甘露醇和高葡萄糖对UHRF2的表达位置没有影响,其表达在细胞质中。
采用划痕实验确定UHRF2是否影响细胞迁移,进而影响细胞上皮间充质转化。结果表明,高糖能够加速细胞迁移(图1c~e),在完全培养基和高糖培养基(不含胎牛血清)中敲低UHRF2可降低细胞迁移率和伤口愈合率。当UHRF2过表达时,细胞迁移和伤口修复加速。上述结果表明UHRF2可能通过影响细胞迁移而影响细胞上皮间充质转化,并在糖尿病肾病纤维化过程中发挥作用。
2.2 高糖促进UBE2V1在HKC中的表达并引发EMT
采用qPCR探究高糖处理细胞的UBE2V1 mRNA表达水平变化,结果如图2a所示。使用甘露醇处理细胞时,对照组和处理组UBE2V1的mRNA表达量无明显差异;但使用高糖处理细胞后,处理组UBE2V1的mRNA表达量明显高于对照组,且在1 h和2 h时具有明显差异。
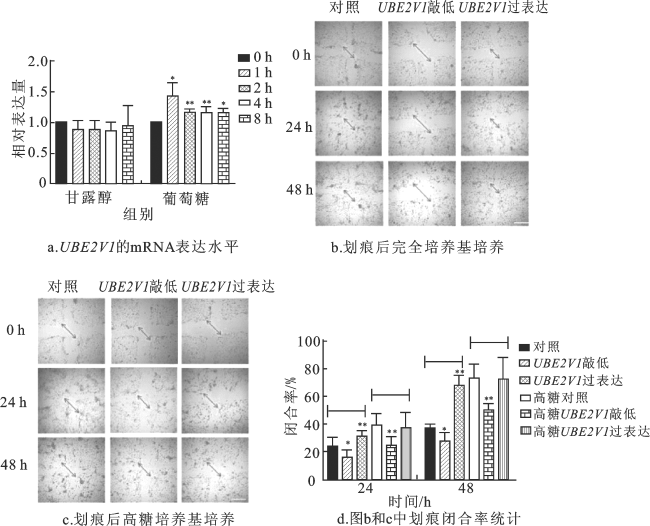
划痕实验结果表明,高糖能够增加细胞迁移(图2b~d)。在完全培养基和高糖培养基中,敲低UBE2V1可显著降低细胞迁移率;在完全培养基中过表达UBE2V1,细胞迁移率显著提高;过表达UBE2V1对高糖环境下的细胞迁移率没有明显影响。这表明UBE2V1可能影响细胞迁移,进而影响上皮间充质转化,并在糖尿病肾病纤维化过程中发挥作用。
2.3 UHRF2和UBE2V1对EMT标志蛋白表达的影响

2.4 UHRF2和UBE2V1协同参与HKC的EMT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 讨论
本研究以HKC为模型,探讨了UHRF2和UBE2V1在高糖培养的HKC中对肾病小管间质纤维化及上皮细胞向间充质细胞转化(EMT)过程的影响,并初步研究了二者在这一过程中的关系。高糖培养提高了UHRF2在HKC中的表达水平,划痕实验显示过表达UHRF2可加速细胞迁移。免疫染色和Western Blot印迹分析表明,在高糖情况下敲低UHRF2可减轻高糖引起的EMT标志蛋白的变化。因此,UHRF2可能对EMT有影响,进而影响糖尿病肾病纤维化。此外,UHRF2完全在细胞质中表达,不受培养条件(甘露醇或高葡萄糖)的影响。
与正常肾脏相比,糖尿病肾病患者的肾脏中UBE2V1的表达量增加[16]。本实验中高糖环境已被证实可增加UBE2V1在HKC中的表达,划痕实验结果显示过表达UBE2V1可加速细胞迁移。免疫染色和Western Blot印迹分析表明,在高糖环境下敲低UBE2V1可以缓解高糖引起的EMT标志蛋白的变化。因此,UBE2V1也可能会影响EMT,进而影响糖尿病肾病纤维化过程。
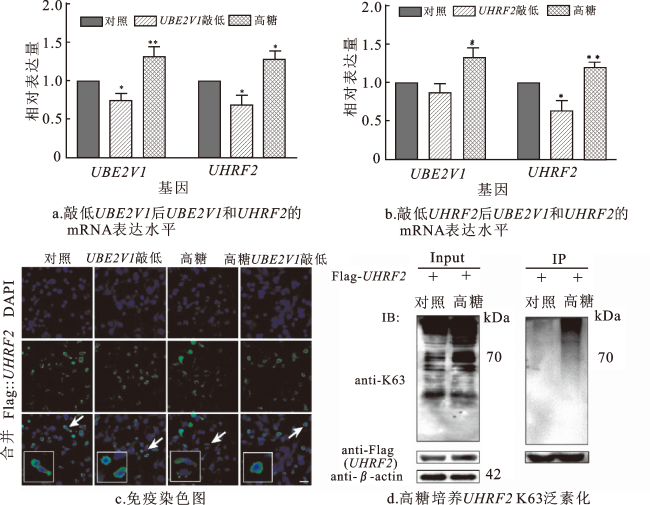
qPCR结果表明,UBE2V1的表达变化会影响UHRF2的表达,但UHRF2对UBE2V1的表达没有影响,反映出UBE2V1和UHRF2可能以协同方式参与EMT,二者可能存在相互作用,UBE2V1可能存在于UHRF2的上游。在免疫染色实验中敲低UBE2V1对UHRF2的表达位置没有影响,UHRF2仍在细胞质中进行表达。免疫沉淀实验表明,高糖条件可增加HKC细胞蛋白的K63泛素化水平,并启动UHRF2的K63泛素化,UBE2V1可以介导高糖培养基中HKC的UHRF2转录上调。因此,UBE2V1可能促进UHRF2的K63泛素化,从而参与EMT加速糖尿病肾病纤维化过程,但其确切的机制还有待进一步研究。
糖尿病肾病是糖尿病的一种严重微血管并发症,已成为终末期肾病的主要病因[25-26],严重威胁着人类的生命安全。目前,糖尿病肾病的治疗方法针对性不强,只是延缓其发病进程,对其发病机制的研究有助于为糖尿病肾病的治疗提供新方向。当前关于UHRF2的研究主要集中在其在DNA甲基化维持和肿瘤中的作用[27-28],本研究结果证实,UHRF2参与了EMT,从而导致了糖尿病肾病纤维化。当细胞被高葡萄糖处理时,UHRF2的K63泛素化水平增加,说明UHRF2可能通过K63泛素化途径参与糖尿病肾病纤维化。同时,UBE2V1可能介导高糖培养HKC中UHRF2的转录上调,并可能促进UHRF2 K63泛素化,从而参与糖尿病肾病纤维化过程。未来可以重点阐明其具体机制,研究敲低UBE2V1对UHRF2 K63泛素化水平的具体影响,这意味着UBE2V1可以促进UHRF2的K63泛素化,从而影响糖尿病肾病纤维化。
4 结论
UHRF2和UBE2V1都会促进高糖环境下肾小管上皮细胞的上皮间充质转化,且UHRF2和UBE2V1可能协同作用促进糖尿病肾病纤维化。研究结果为糖尿病肾病的发病机制提供了理论依据,并为今后利用小鼠模型进行糖尿病肾病研究奠定了基础。目前,关于UHRF2在糖尿病肾病中作用的研究未见报道,本研究为糖尿病肾病的预防和治疗提供了新的思路和方向。